

A mutational analysis of U12-dependent splice site dinucleotides
- PMID: 16043500
- PMCID: PMC1370826
- DOI: 10.1261/rna.7206305
A mutational analysis of U12-dependent splice site dinucleotides
Abstract
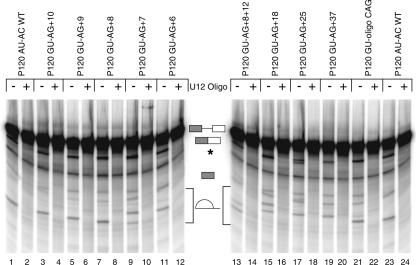
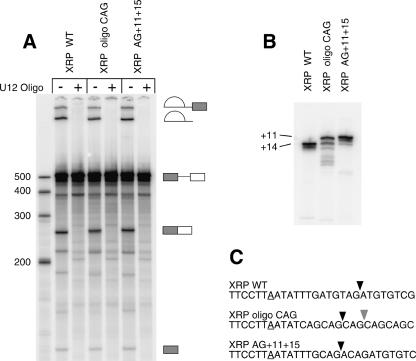
Introns spliced by the U12-dependent minor spliceosome are divided into two classes based on their splice site dinucleotides. The /AU-AC/ class accounts for about one-third of U12-dependent introns in humans, while the /GU-AG/ class accounts for the other two-thirds. We have investigated the in vivo and in vitro splicing phenotypes of mutations in these dinucleotide sequences. A 5' A residue can splice to any 3' residue, although C is preferred. A 5' G residue can splice to 3' G or U residues with a preference for G. Little or no splicing was observed to 3' A or C residues. A 5' U or C residue is highly deleterious for U12-dependent splicing, although some combinations, notably 5' U to 3' U produced detectable spliced products. The dependence of 3' splice site activity on the identity of the 5' residue provides evidence for communication between the first and last nucleotides of the intron. Most mutants in the second position of the 5' splice site and the next to last position of the 3' splice site were defective for splicing. Double mutants of these residues showed no evidence of communication between these nucleotides. Varying the distance between the branch site and the 3' splice site dinucleotide in the /GU-AG/ class showed that a somewhat larger range of distances was functional than for the /AU-AC/ class. The optimum branch site to 3' splice site distance of 11-12 nucleotides appears to be the same for both classes.
Figures
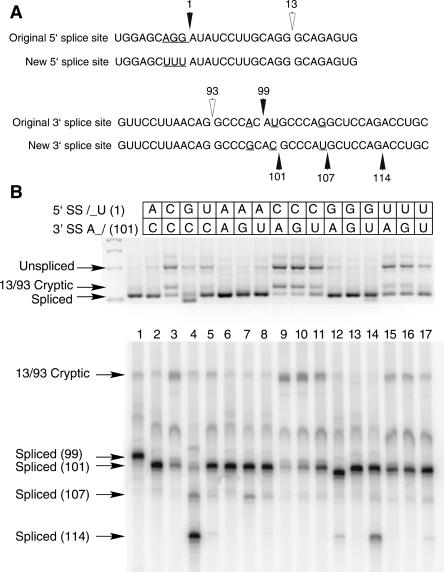
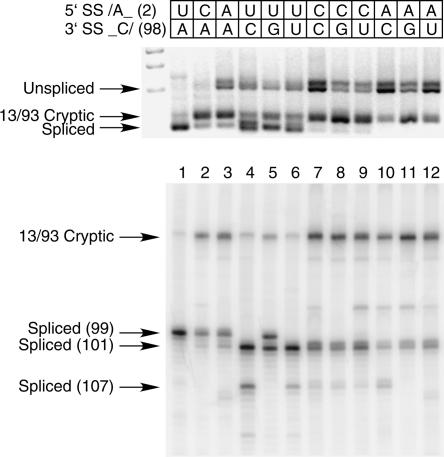
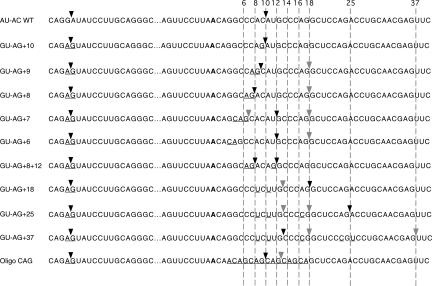
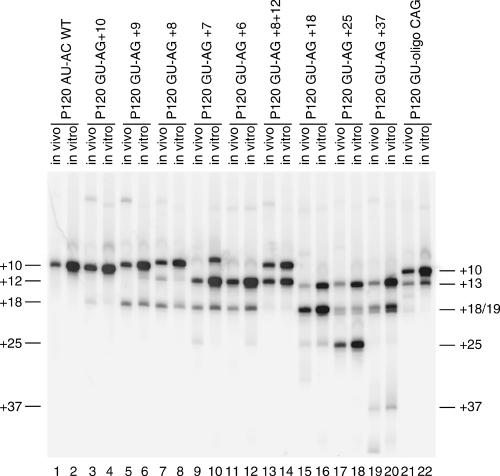
References
-
- Aebi, M., Hornig, H., Padgett, R.A., Reiser, J., and Weissmann, C. 1986. Sequence requirements for splicing of higher eukaryotic nuclear pre-mRNA. Cell 47: 555–565. - PubMed
-
- Burge, C.B. and Sharp, P.A. 1997. Classification of introns: U2-type or U12-type. Cell 91: 875–879. - PubMed
-
- Burge, C.B., Tuschl, T., and Sharp, P.A. 1999. Splicing of precursors to mRNAs by the spliceosome. In The RNA world, 2d ed. (eds. R.F. Gestland et al.), pp. 525–560. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources