

Probing the geometry of the inner vestibule of BK channels with sugars
- PMID: 16043773
- PMCID: PMC2266569
- DOI: 10.1085/jgp.200509286
Probing the geometry of the inner vestibule of BK channels with sugars
Abstract
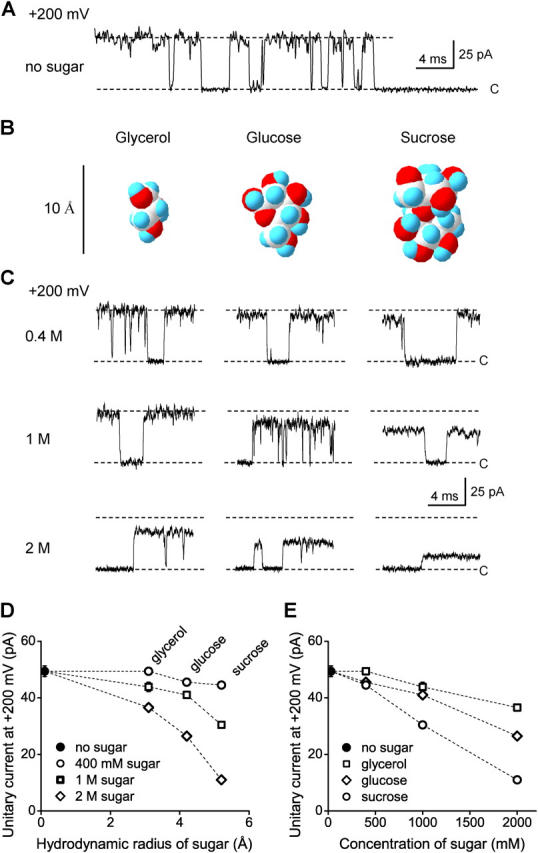
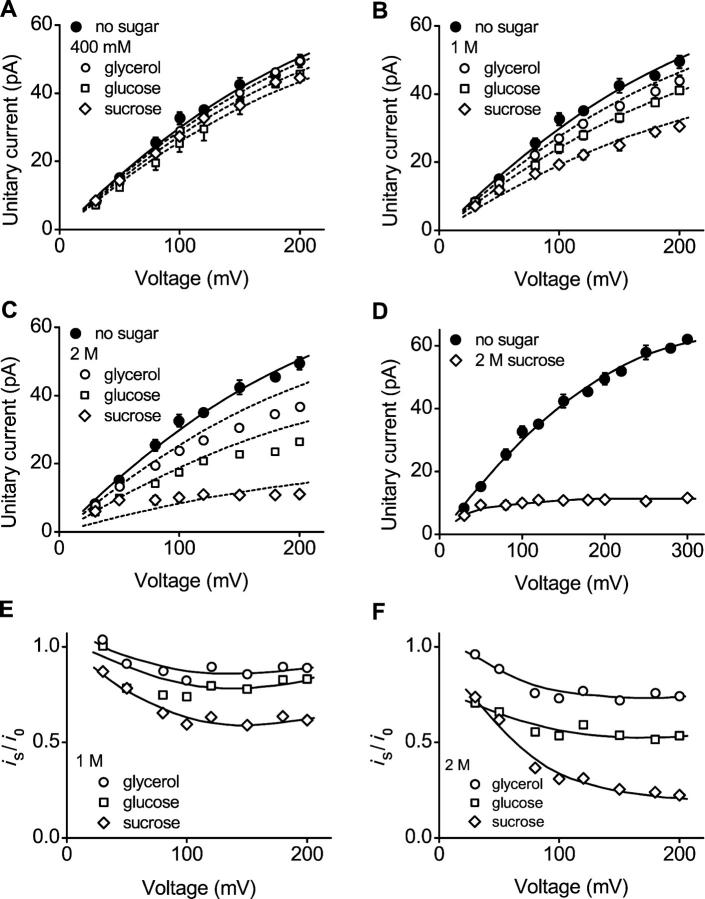
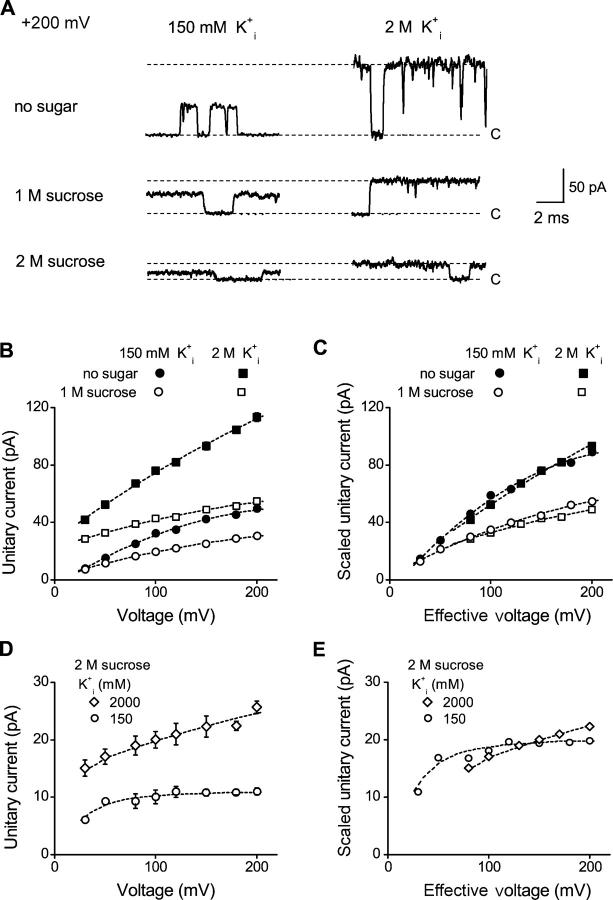
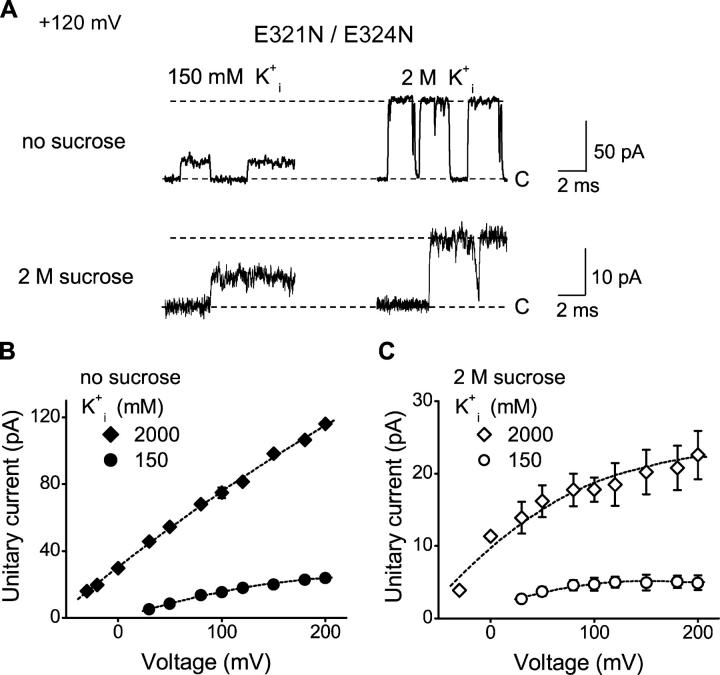
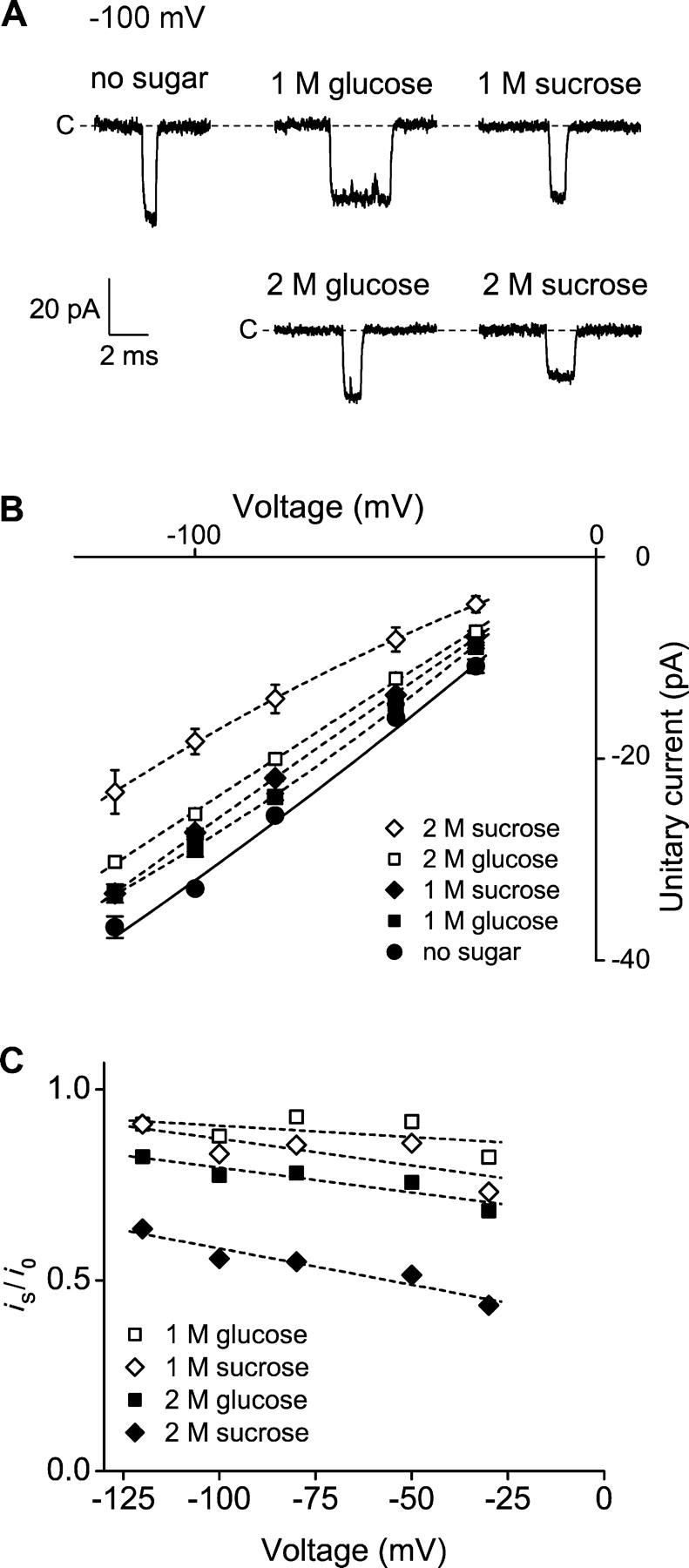
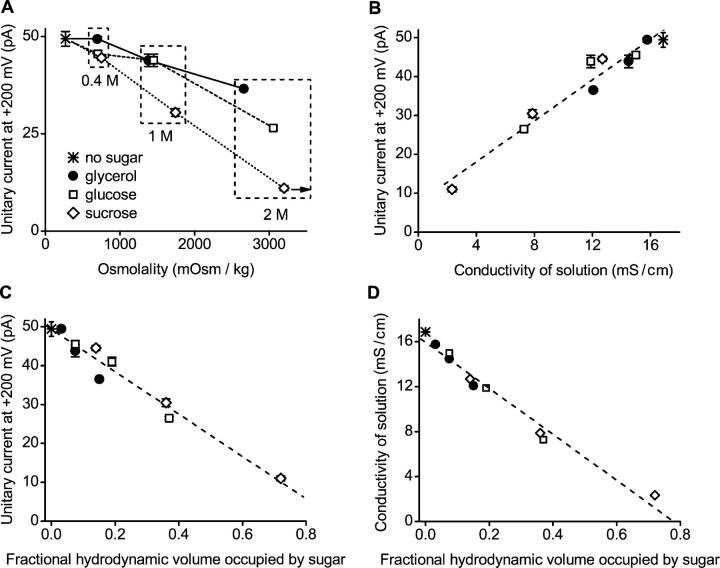
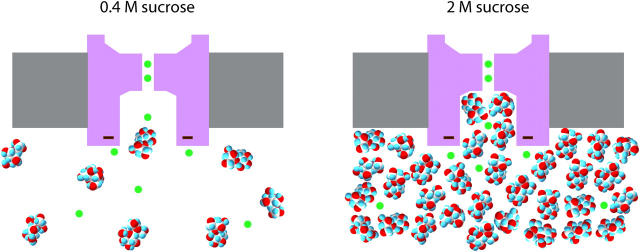
The geometry of the inner vestibule of BK channels was probed by examining the effects of different sugars in the intracellular solution on single-channel current amplitude (unitary current). Glycerol, glucose, and sucrose decreased unitary current through BK channels in a concentration- and size-dependent manner, in the order sucrose > glucose > glycerol, with outward currents being reduced more than inward currents. The fractional decrease of outward current was more directly related to the fractional hydrodynamic volume occupied by the sugars than to changes in osmolality. For concentrations of sugars < or =1 M, the i/V plots for outward currents in the presence and absence of sugar superimposed after scaling, and increasing K(+)(i) from 150 mM to 2 M increased the magnitudes of the i/V plots with little effect on the shape of the scaled curves. These observations suggest that sugars < or =1 M reduce outward currents mainly by entering the inner vestibule and reducing the movement of K(+) through the vestibule, rather than by limiting diffusion-controlled access of K(+) to the vestibule. With 2 M sucrose, the movement of K(+) into the inner vestibule became diffusion limited for 150 mM K(+)(i) and voltages > +100 mV. Increasing K(+)(i) then relieved the diffusion limitation. An estimate of the capture radius based on the 5 pA diffusion-limited current for channels without the ring of negative charge at the entrance to the inner vestibule was 2.2 A. Adding the radius of a hydrated K(+) (6-8 A) then gave an effective radius for the entrance to the inner vestibule of 8-10 A. Such a functionally wide entrance to the inner vestibule together with our observation that even small concentrations of sugar in the inner vestibule reduce unitary current suggest that a wide inner vestibule is required for the large conductance of BK channels.
Figures
References
-
- Arakawa, T., and S.N. Timasheff. 1982. Stabilization of protein structure by sugars. Biochemistry. 21:6536–6544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
