

Dynamic control of lymphocyte trafficking by fever-range thermal stress
- PMID: 16044255
- PMCID: PMC11030888
- DOI: 10.1007/s00262-005-0022-9
Dynamic control of lymphocyte trafficking by fever-range thermal stress
Abstract
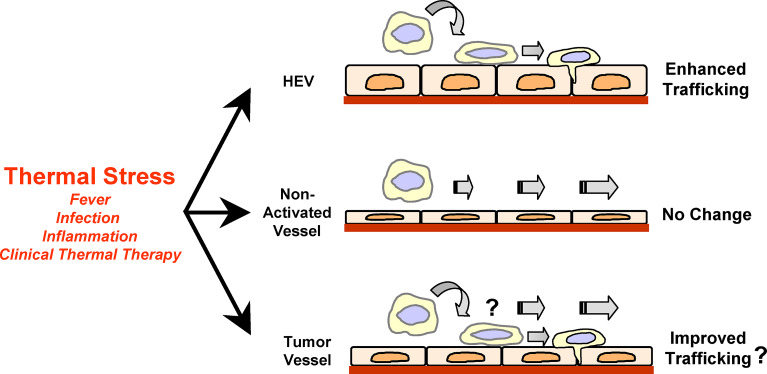
Migration of blood-borne lymphocytes into tissues involves a tightly orchestrated sequence of adhesion events. Adhesion molecules and chemokine receptors on the surface of circulating lymphocytes initiate contact with specialized endothelial cells under hemodynamic shear prior to extravasation across the vascular barrier into tissues. Lymphocyte-endothelial adhesion occurs preferentially in high endothelial venules (HEV) of peripheral lymphoid organs. The continuous recirculation of naïve and central memory lymphocytes across lymph node and Peyer's patch HEV underlies immune surveillance and immune homeostasis. Lymphocyte-endothelial interactions are markedly enhanced in HEV-like vessels of extralymphoid organs during physiological responses associated with acute and chronic inflammation. Similar adhesive mechanisms must be invoked for efficient trafficking of immune effector cells to tumor sites in order for the immune system to have an impact on tumor progression. Here we discuss recent evidence for the role of fever-range thermal stress in promoting lymphocyte-endothelial adhesion and trafficking across HEV in peripheral lymphoid organs. Findings are also presented that support the hypothesis that lymphocyte-endothelial interactions are limited within tumor microenvironments. Further understanding of the molecular mechanisms that dynamically promote lymphocyte trafficking in HEV may provide the basis for novel approaches to improve recruitment of immune effector cells to tumor sites.
Figures
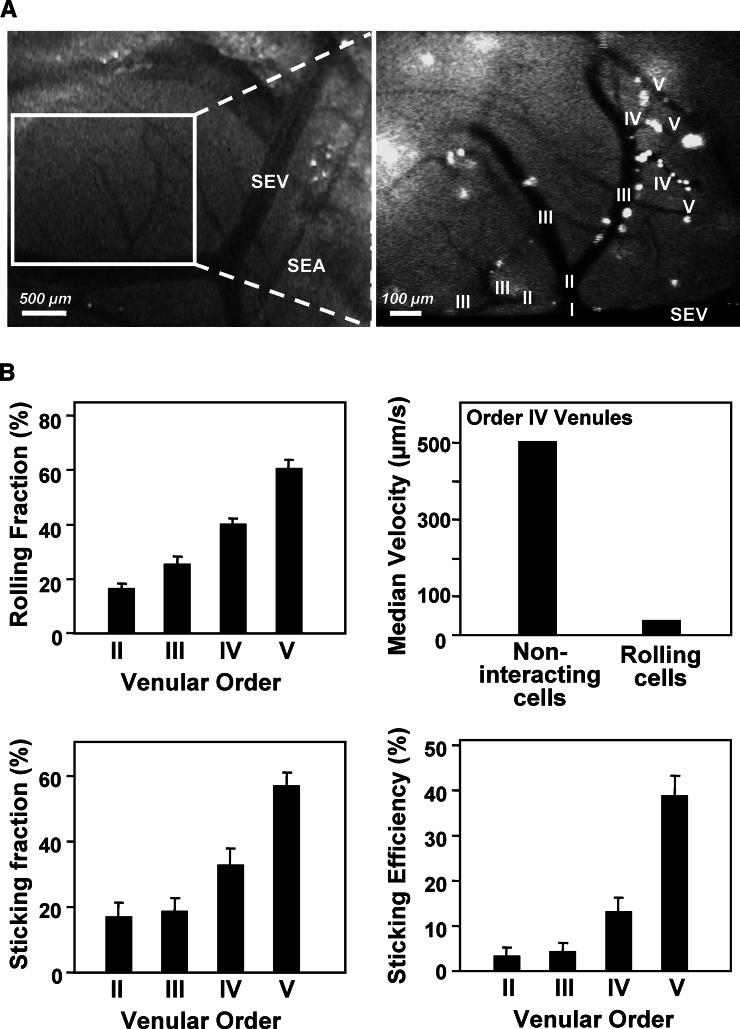
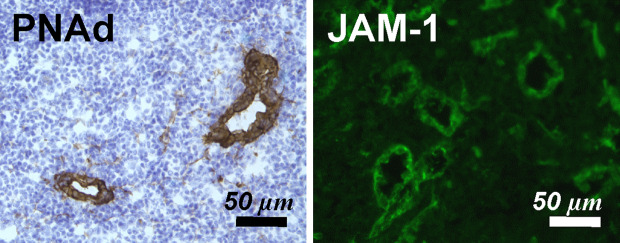
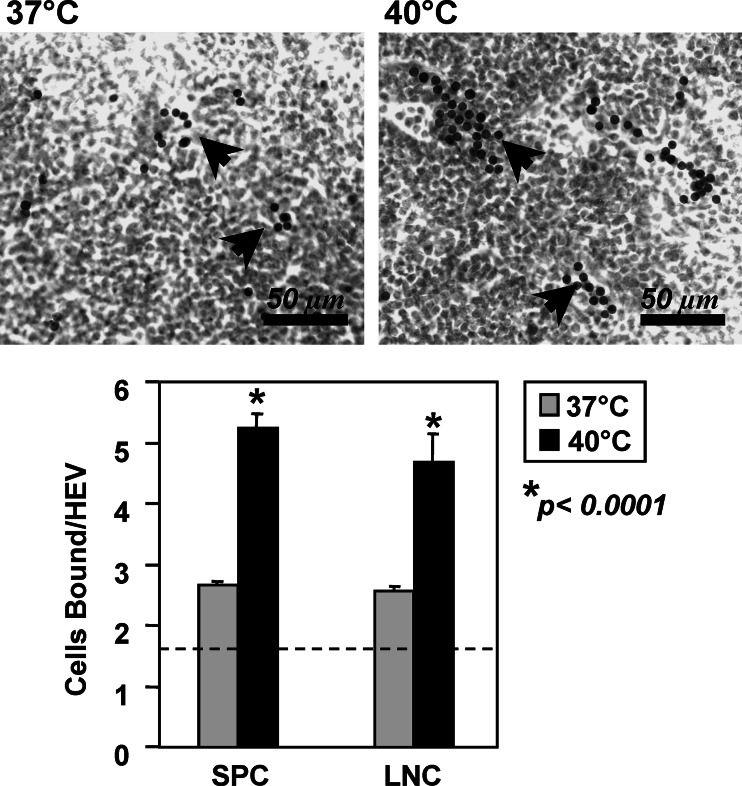
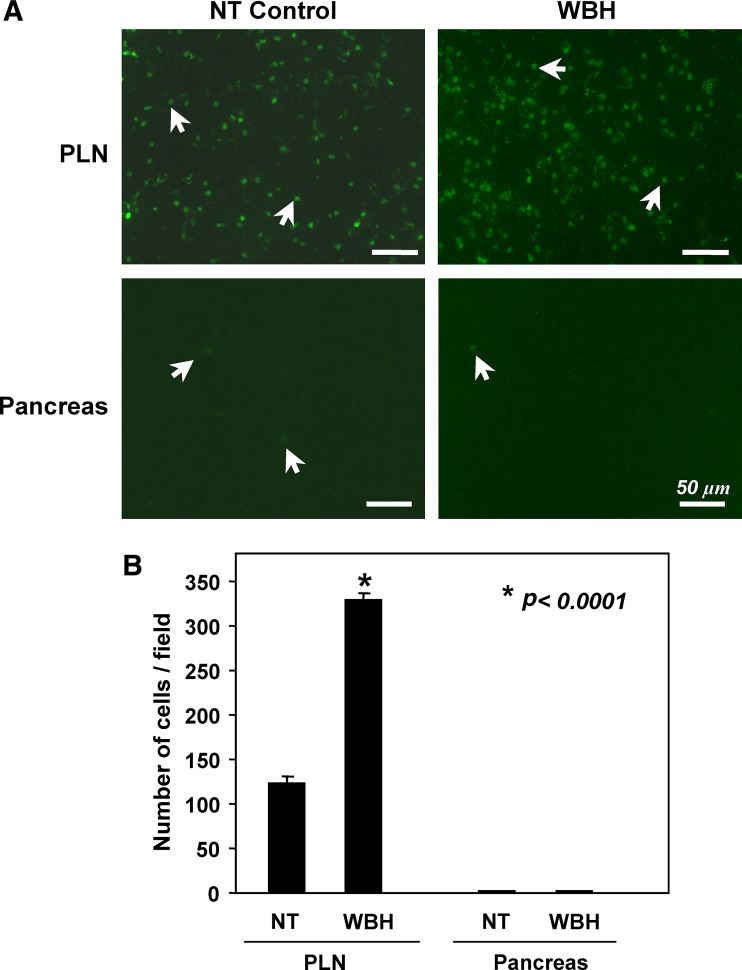
Similar articles
-
Fever-range thermal stress promotes lymphocyte trafficking across high endothelial venules via an interleukin 6 trans-signaling mechanism.Nat Immunol. 2006 Dec;7(12):1299-308. doi: 10.1038/ni1406. Nat Immunol. 2006. PMID: 17086187
-
Targeted regulation of a lymphocyte-endothelial-interleukin-6 axis by thermal stress.Int J Hyperthermia. 2008 Feb;24(1):67-78. doi: 10.1080/02656730701772498. Int J Hyperthermia. 2008. PMID: 18214770 Review.
-
Fever-range hyperthermia dynamically regulates lymphocyte delivery to high endothelial venules.Blood. 2001 May 1;97(9):2727-33. doi: 10.1182/blood.v97.9.2727. Blood. 2001. PMID: 11313264
-
Fine-tuning immune surveillance by fever-range thermal stress.Immunol Res. 2010 Mar;46(1-3):177-88. doi: 10.1007/s12026-009-8122-9. Immunol Res. 2010. PMID: 19760057 Free PMC article. Review.
-
Hurdles to lymphocyte trafficking in the tumor microenvironment: implications for effective immunotherapy.Immunol Invest. 2006;35(3-4):251-77. doi: 10.1080/08820130600745430. Immunol Invest. 2006. PMID: 16916754 Review.
Cited by
-
Sublethal laser interstitial thermal therapy leads to confined neurological deficits and glioblastoma regression.Lasers Med Sci. 2025 Jul 12;40(1):312. doi: 10.1007/s10103-025-04568-8. Lasers Med Sci. 2025. PMID: 40646361
-
Hyperthermic intraperitoneal chemotherapy and colorectal cancer: From physiology to surgery.World J Clin Cases. 2022 Oct 26;10(30):10852-10861. doi: 10.12998/wjcc.v10.i30.10852. World J Clin Cases. 2022. PMID: 36338235 Free PMC article. Review.
-
Regulation of a lymphocyte-endothelial-IL-6 trans-signaling axis by fever-range thermal stress: hot spot of immune surveillance.Cytokine. 2007 Jul;39(1):84-96. doi: 10.1016/j.cyto.2007.07.184. Cytokine. 2007. PMID: 17903700 Free PMC article. Review.
-
Crosstalk of Inflammatory Cytokines within the Breast Tumor Microenvironment.Int J Mol Sci. 2023 Feb 16;24(4):4002. doi: 10.3390/ijms24044002. Int J Mol Sci. 2023. PMID: 36835413 Free PMC article. Review.
-
IL-17 promotes neutrophil entry into tumor-draining lymph nodes following induction of sterile inflammation.J Immunol. 2013 Oct 15;191(8):4348-57. doi: 10.4049/jimmunol.1103621. Epub 2013 Sep 11. J Immunol. 2013. PMID: 24026079 Free PMC article.
References
-
- Butcher EC, Williams M, Youngman K, Rott L, Briskin M. Lymphocyte trafficking and regional immunity. Adv Immunol. 1999;72:209–253. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
