

Programmed and induced phenotype of the hippocampal granule cells
- PMID: 16049169
- PMCID: PMC6724843
- DOI: 10.1523/JNEUROSCI.1674-05.2005
Programmed and induced phenotype of the hippocampal granule cells
Abstract
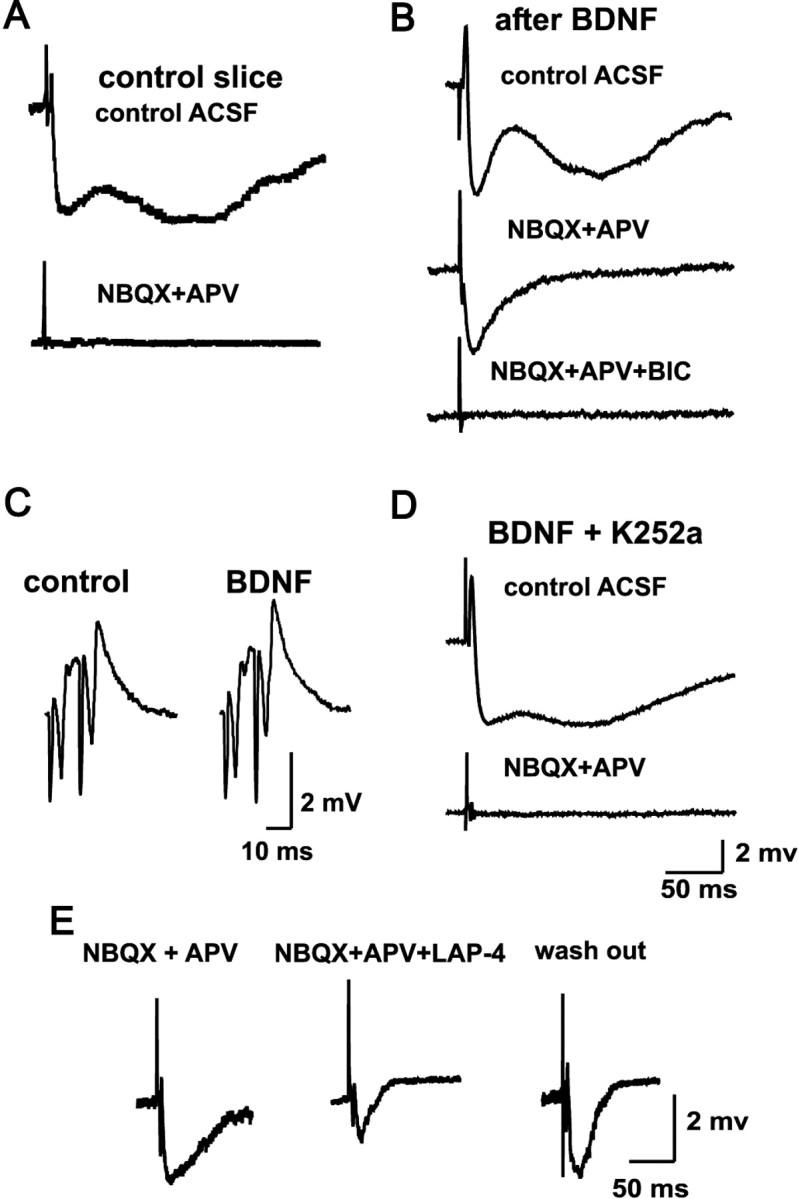
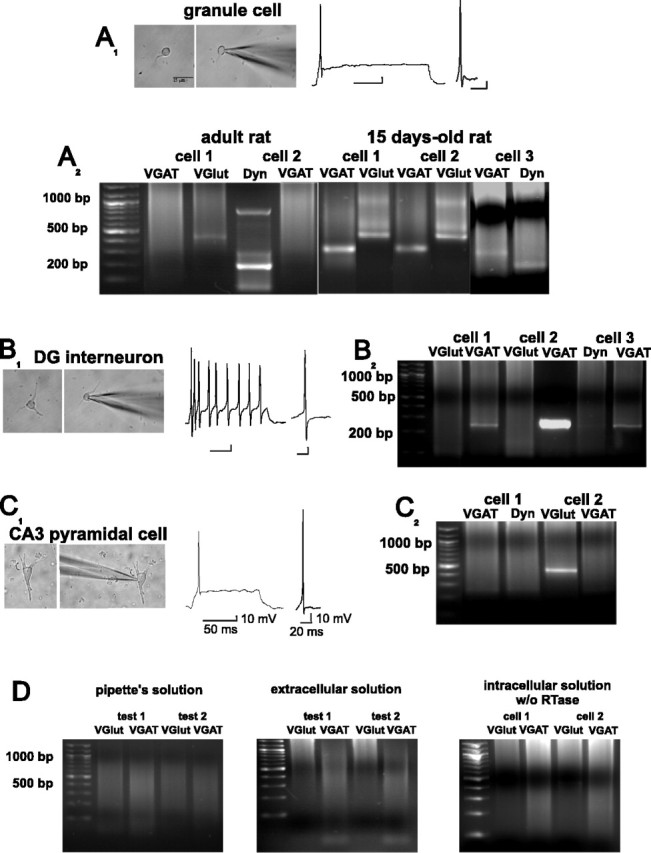
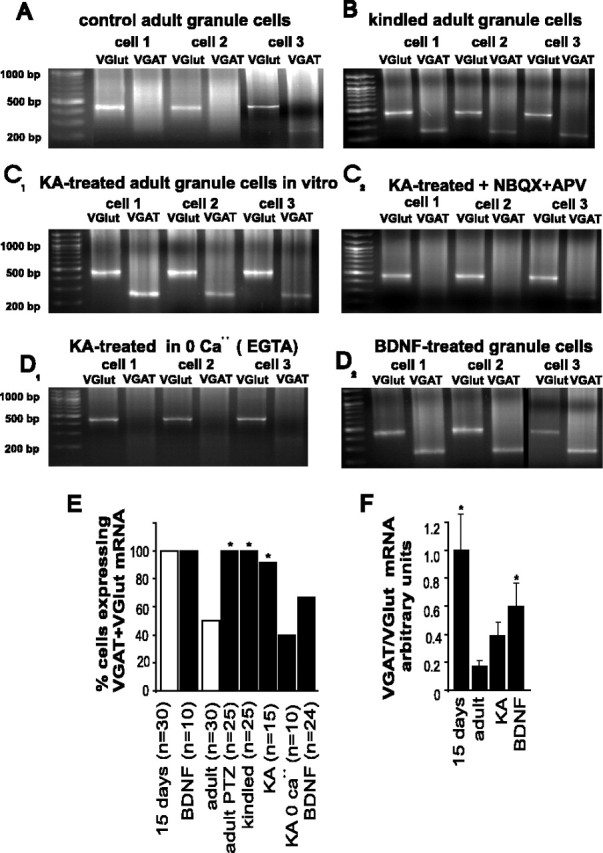
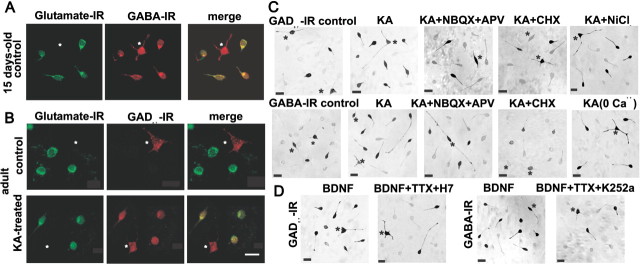
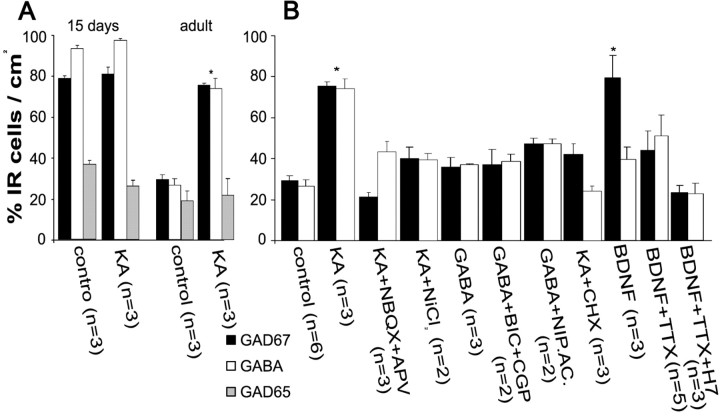
Certain neurons choose the neurotransmitter they use in an activity-dependent manner, and trophic factors are involved in this phenotypic differentiation during development. Developing hippocampal granule cells (GCs) constitutively express the markers of the glutamatergic and GABAergic phenotypes, but when development is completed, the GABAergic phenotype shuts off. With electrophysiological, single-cell reverse transcription-PCR and immunohistological techniques, we show here that short-term (24 h) cultures of fully differentiated adult glutamatergic GCs, which express glutamate, VGlut-1 (vesicular glutamate transporter) mRNA, calbindin, and dynorphin mRNA, can be induced to reexpress the GABAergic markers GABA, GAD67 (glutamate decarboxylase 67 kDa isoform), and VGAT (vesicular GABA transporter) mRNA, by sustained synaptic or direct activation of glutamate receptors and by activation of TrkB (tyrosine receptor kinase B) receptors, with brain-derived neurotrophic factor (BDNF) (30 min). The expression of the GABAergic markers was prevented by the blockade of glutamate receptors and sodium or calcium channels, and by inhibitors of protein kinases and protein synthesis. In hippocampal slices of epileptic rats and in BDNF-treated slices from naive rats, we confirmed the appearance of monosynaptic GABAA receptor-mediated responses to GC stimulation, in the presence of glutamate receptors blockers. Accordingly, GC cultures prepared from these slices showed the coexpression of the glutamatergic and GABAergic markers. Our results demonstrate that the neurotransmitter choice of the GCs, which are unique in terms of their continuing birth and death throughout life, depends on programmed and environmental factors, and this process is neither limited by a critical developmental period nor restricted by their insertion in their natural network.
Figures
References
-
- Altar CA, DiStefano PS (1998) Neurotrophin trafficking by anterograde transport. Trends Neurosci 21: 433-437. - PubMed
-
- Ben-Ari Y, Tseeb V, Raggozzino D, Khazipov R, Gaïarsa JL (1994) gamma-Aminobutyric acid (GABA): a fast excitatory transmitter which may regulate the development of hippocampal neurones in early postnatal life. Prog Brain Res 102: 261-273. - PubMed
-
- Bergersen L, Ruiz A, Bjaalie JG, Kullmann DM, Gundersen V (2003) GABA and GABAA receptors at hippocampal mossy fibre synapses. Eur J Neurosci 18: 931-941. - PubMed
-
- Berninger B, Garcia DE, Inagaki N, Hahnel C, Lindholm D (1993) BDNF and NT-3 induce intracellular Ca2+ elevation in hippocampal neurones. NeuroReport 4: 1303-1306. - PubMed
-
- Berzaghi MP, Gutiérrez R, Heinemann U, Lindholm D, Thoenen H (1995) Neurotrophins induce acute transmitter-mediated changes in brain electrical activity. Soc Neurosci Abstr 21: 226.3.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous