

Oscillatory bursts in the optic tectum of birds represent re-entrant signals from the nucleus isthmi pars parvocellularis
- PMID: 16049185
- PMCID: PMC6724834
- DOI: 10.1523/JNEUROSCI.1379-05.2005
Oscillatory bursts in the optic tectum of birds represent re-entrant signals from the nucleus isthmi pars parvocellularis
Erratum in
- J Neurosci. 2005 Aug 3;25(31):table of contents. Mpdozis, Jorge [corrected to Mpodozis, Jorge]
Abstract
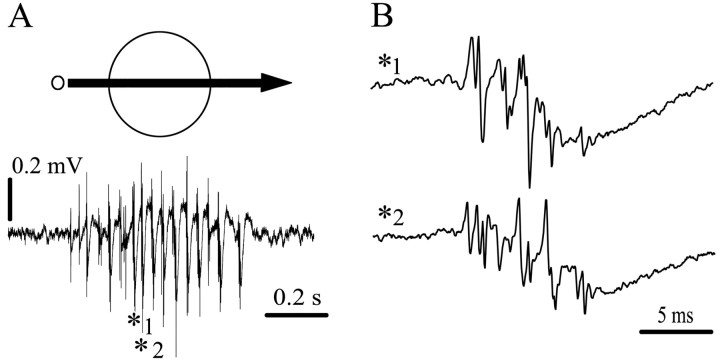
Fast oscillatory bursts (OBs; 500-600 Hz) are the most prominent response to visual stimulation in the optic tectum of birds. To investigate the neural mechanisms generating tectal OBs, we compared local recordings of OBs with simultaneous intracellular and extracellular single-unit recordings in the tectum of anesthetized pigeons. We found a specific population of units that responded with burst discharges that mirrored the burst pattern of OBs. Intracellular filling with biocytin of some of these bursting units demonstrated that they corresponded to the paintbrush axon terminals from the nucleus isthmi pars parvocellularis (Ipc). Direct recordings in the Ipc confirmed the high correlation between Ipc cell firing and tectal OBs. After injecting micro-drops of lidocaine in the Ipc, the OBs of the corresponding tectal locus disappeared completely. These results identify the paintbrush terminals as the neural elements generating tectal OBs. These terminals are presumably cholinergic and ramify across tectal layers in a columnar manner. Because the optic tectum and the Ipc are reciprocally connected such that each Ipc neuron sends a paintbrush axon to the part of the optic tectum from which its visual inputs come, tectal OBs represent re-entrant signals from the Ipc, and the spatial-temporal pattern of OBs across the tectum is the mirror representation of the spatial-temporal pattern of bursting neurons in the Ipc. We propose that an active location in the Ipc may act, via bursting paintbrushes in the tectum, as a focal "beam of attention" across tectal layers, enhancing the saliency of stimuli in the corresponding location in visual space.
Figures






Similar articles
-
A cholinergic gating mechanism controlled by competitive interactions in the optic tectum of the pigeon.J Neurosci. 2007 Jul 25;27(30):8112-21. doi: 10.1523/JNEUROSCI.1420-07.2007. J Neurosci. 2007. PMID: 17652602 Free PMC article.
-
A blinking focal pattern of re-entrant activity in the avian tectum.Curr Biol. 2023 Jan 9;33(1):1-14.e4. doi: 10.1016/j.cub.2022.10.070. Epub 2022 Nov 28. Curr Biol. 2023. PMID: 36446352
-
Axon terminals from the nucleus isthmi pars parvocellularis control the ascending retinotectofugal output through direct synaptic contact with tectal ganglion cell dendrites.J Comp Neurol. 2016 Feb 1;524(2):362-79. doi: 10.1002/cne.23860. Epub 2015 Aug 18. J Comp Neurol. 2016. PMID: 26224333
-
The nucleus isthmi and dual modulation of the receptive field of tectal neurons in non-mammals.Brain Res Brain Res Rev. 2003 Jan;41(1):13-25. doi: 10.1016/s0165-0173(02)00217-5. Brain Res Brain Res Rev. 2003. PMID: 12505645 Review.
-
Functional anatomy of the tectum mesencephali of the goldfish. An explorative analysis of the functional implications of the laminar structural organization of the tectum.Brain Res. 1983 Dec;287(3):247-97. doi: 10.1016/0165-0173(83)90008-5. Brain Res. 1983. PMID: 6362772 Review.
Cited by
-
Visual categories and concepts in the avian brain.Anim Cogn. 2023 Jan;26(1):153-173. doi: 10.1007/s10071-022-01711-8. Epub 2022 Nov 10. Anim Cogn. 2023. PMID: 36352174 Free PMC article. Review.
-
Mechanisms of competitive selection: A canonical neural circuit framework.Elife. 2020 May 20;9:e51473. doi: 10.7554/eLife.51473. Elife. 2020. PMID: 32431293 Free PMC article.
-
Resonant Cholinergic Dynamics in Cognitive and Motor Decision-Making: Attention, Category Learning, and Choice in Neocortex, Superior Colliculus, and Optic Tectum.Front Neurosci. 2016 Jan 20;9:501. doi: 10.3389/fnins.2015.00501. eCollection 2015. Front Neurosci. 2016. PMID: 26834535 Free PMC article.
-
Donut-like organization of inhibition underlies categorical neural responses in the midbrain.Nat Commun. 2022 Mar 30;13(1):1680. doi: 10.1038/s41467-022-29318-0. Nat Commun. 2022. PMID: 35354821 Free PMC article.
-
Stimulus-specific adaptations in the gaze control system of the barn owl.J Neurosci. 2008 Feb 6;28(6):1523-33. doi: 10.1523/JNEUROSCI.3785-07.2008. J Neurosci. 2008. PMID: 18256273 Free PMC article.
References
-
- Bagnoli P, Fontanesi G, Alesci R, Erichsen J (1992) Distribution of neuropeptide Y, substance P, and choline acetyltransferase in the developing visual system of the pigeon and effects of unilateral retina removal. J Comp Neurol 318: 392-414. - PubMed
-
- Baizer JS, Whitney JF, Bender DB (1991) Bilateral projections from the parabigeminal nucleus to the superior colliculus in monkey. Exp Brain Res 86: 467-470. - PubMed
-
- Binns KE, Salt TE (2000) The functional influence of nicotinic cholinergic receptors on the visual responses of neurons in the superficial superior colliculus. Vis Neurosci 17: 283-289. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous