

RNAs in the virion of Kaposi's sarcoma-associated herpesvirus
- PMID: 16051806
- PMCID: PMC1182685
- DOI: 10.1128/JVI.79.16.10138-10146.2005
RNAs in the virion of Kaposi's sarcoma-associated herpesvirus
Abstract
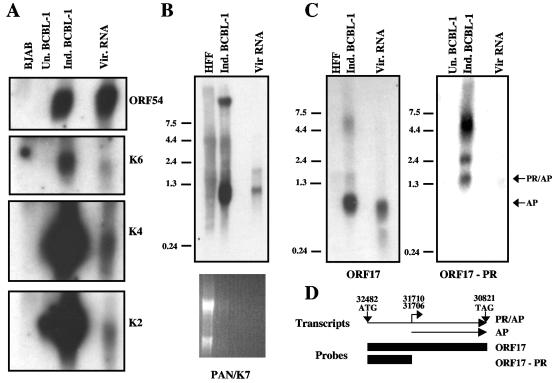
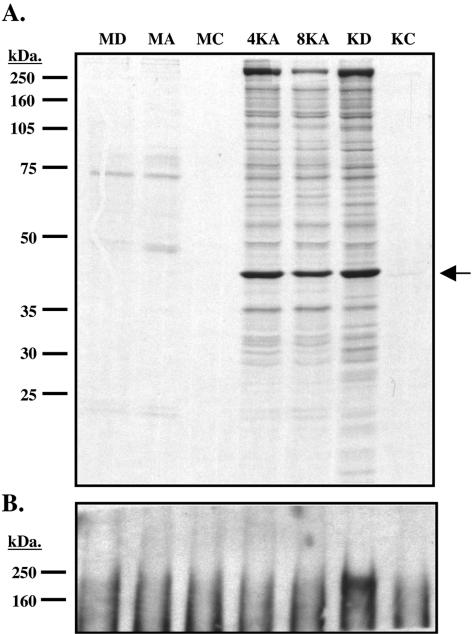
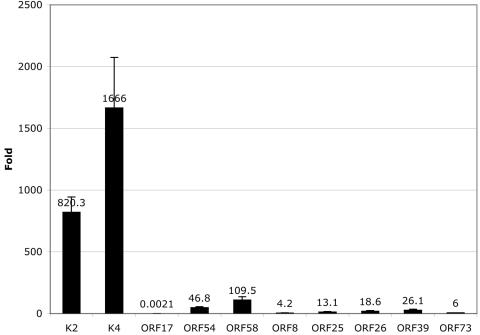
De novo infection of cultured cells with Kaposi's sarcoma-associated herpesvirus (KSHV) typically results in a latent infection. Recently, however, it has been reported that a subset of lytic mRNAs can be detected in cells shortly after KSHV infection; this expression is transient and eventually subsides, leading to latent infection (H. H. Krishnan et al., J. Virol 78:3601-3620, 2004). Since it has been shown that viral RNAs can be packaged into other herpesvirus virions, we sought to determine if KSHV virions contained RNAs and, if so, whether these RNAs contributed to the pool of lytic transcripts detected immediately after infection. Using DNA microarray, reverse transcription (RT)-PCR, and Northern blotting analyses, we identified 11 virally encoded RNAs in KSHV virions. These corresponded in size to the full-length mRNAs found in cytoplasmic RNA, and at least one was directly demonstrated to be translated upon infection in the presence of actinomycin D. Ten of these RNAs correspond to transcripts reported by Krishnan et al. at early times of infection, representing ca. 30% of such RNAs. Thus, import of RNAs in virions can account for some but not all of the early-appearing lytic transcripts. Quantitative RT-PCR analysis of infected-cell RNA demonstrated that most of the virion RNAs were very abundant at late times of infection, consistent with nonspecific incorporation during budding. However, the intracellular levels of one virion mRNA, encoding the viral protease, were much lower than those of transcripts not packaged in the virus particle, strongly suggesting that it may be incorporated by a specific mechanism.
Figures


References
-
- Akula, S. M., N. P. Pramod, F. Z. Wang, and B. Chandran. 2002. Integrin α3β1 (CD 49c/29) is a cellular receptor for Kaposi's sarcoma-associated herpesvirus (KSHV/HHV-8) entry into the target cells. Cell 108:407-419. - PubMed
-
- Ballestas, M. E., P. A. Chatis, and K. M. Kaye. 1999. Efficient persistence of extrachromosomal KSHV DNA mediated by latency-associated nuclear antigen. Science 284:641-644. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
