

Detection of reactive oxygen species in isolated, perfused lungs by electron spin resonance spectroscopy
- PMID: 16053530
- PMCID: PMC1184103
- DOI: 10.1186/1465-9921-6-86
Detection of reactive oxygen species in isolated, perfused lungs by electron spin resonance spectroscopy
Abstract
Background: The sources and measurement of reactive oxygen species (ROS) in intact organs are largely unresolved. This may be related to methodological problems associated with the techniques currently employed for ROS detection. Electron spin resonance (ESR) with spin trapping is a specific method for ROS detection, and may address some these technical problems.
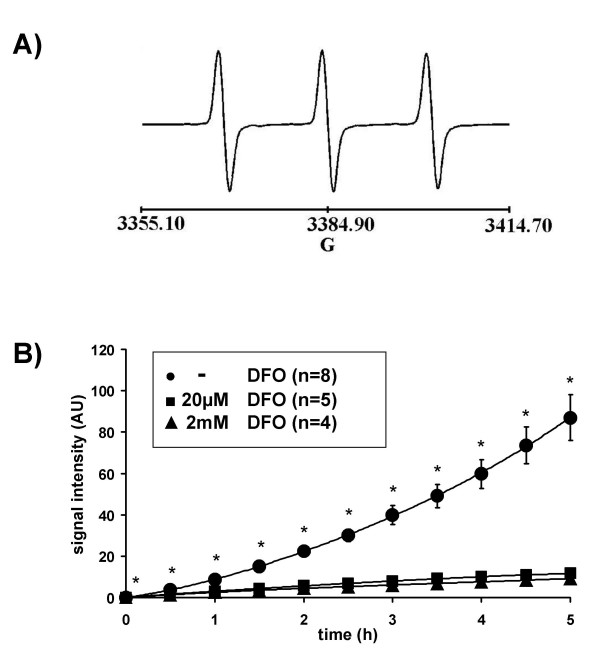
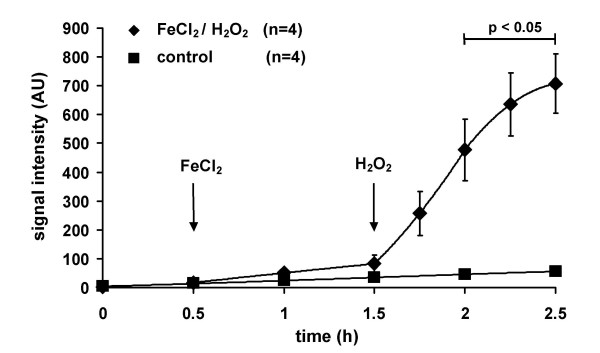
Methods: We have established a protocol for the measurement of intravascular ROS release from isolated buffer-perfused and ventilated rabbit and mouse lungs, combining lung perfusion with the spin probe 1-hydroxy-3-carboxy-2,2,5,5-tetramethylpyrrolidine (CPH) and ESR spectroscopy. We then employed this technique to characterize hypoxia-dependent ROS release, with specific attention paid to NADPH oxidase-dependent superoxide formation as a possible vasoconstrictor pathway.
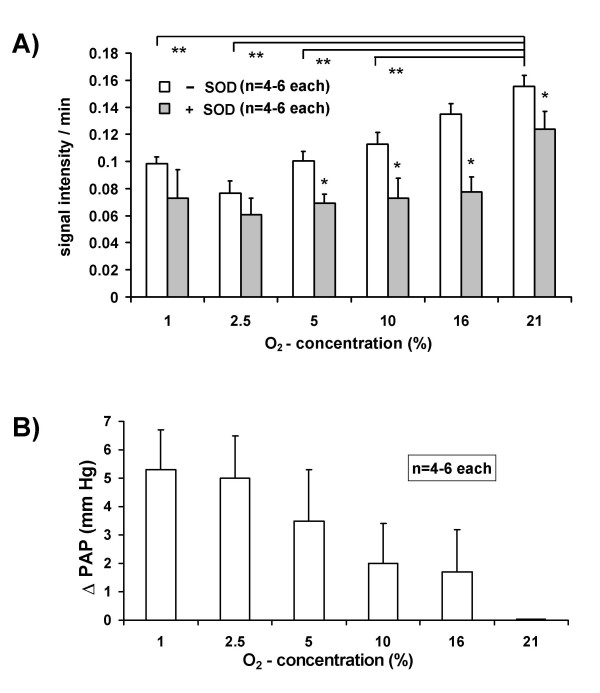
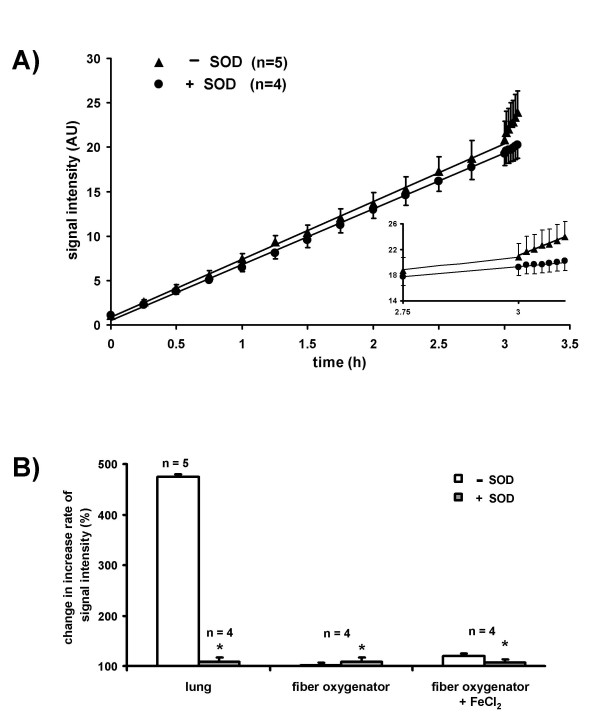
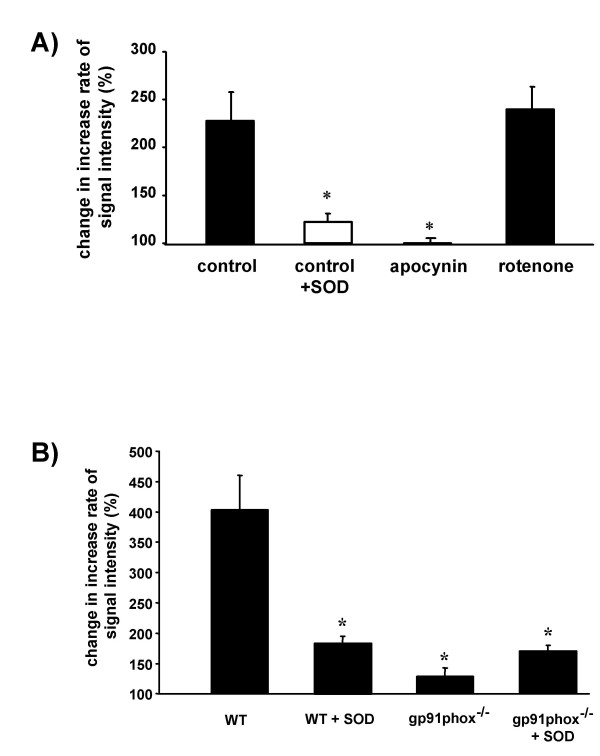
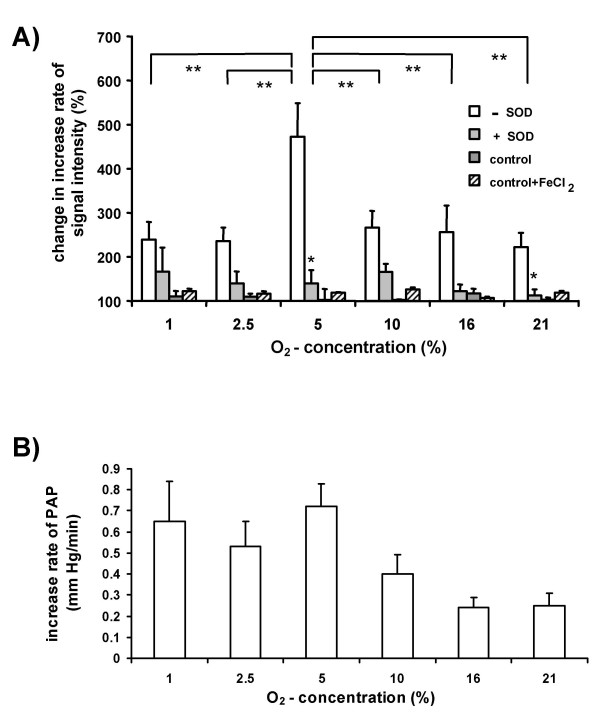
Results: While perfusing lungs with CPH over a range of inspired oxygen concentrations (1-21 %), the rate of CP* formation exhibited an oxygen-dependence, with a minimum at 2.5 % O2. Addition of superoxide dismutase (SOD) to the buffer fluid illustrated that a minor proportion of this intravascular ROS leak was attributable to superoxide. Stimulation of the lungs by injection of phorbol-12-myristate-13-acetate (PMA) into the pulmonary artery caused a rapid increase in CP* formation, concomitant with pulmonary vasoconstriction. Both the PMA-induced CPH oxidation and the vasoconstrictor response were largely suppressed by SOD. When the PMA challenge was performed at different oxygen concentrations, maximum superoxide liberation and pulmonary vasoconstriction occurred at 5% O2. Using a NADPH oxidase inhibitor and NADPH-oxidase deficient mice, we illustrated that the PMA-induced superoxide release was attributable to the stimulation of NADPH oxidases.
Conclusion: The perfusion of isolated lungs with CPH is suitable for detection of intravascular ROS release by ESR spectroscopy. We employed this technique to demonstrate that 1) PMA-induced vasoconstriction is caused "directly" by superoxide generated from NADPH oxidases and 2) this pathway is pronounced in hypoxia. NADPH oxidases thus may contribute to the hypoxia-dependent regulation of pulmonary vascular tone.
Figures
Similar articles
-
Vanadium-induced apoptosis and pulmonary inflammation in mice: Role of reactive oxygen species.J Cell Physiol. 2003 Apr;195(1):99-107. doi: 10.1002/jcp.10232. J Cell Physiol. 2003. PMID: 12599213
-
Superoxide anion, the main species of ROS in the development of ARDS induced by oleic acid.Free Radic Res. 2004 Dec;38(12):1281-7. doi: 10.1080/10715760400006940. Free Radic Res. 2004. PMID: 15763952
-
EPR detection of reactive oxygen species in hemolymph of Galleria mellonella and Dendrolimus superans sibiricus (Lepidoptera) larvae.Biochem Biophys Res Commun. 1999 Oct 14;264(1):212-5. doi: 10.1006/bbrc.1999.1504. Biochem Biophys Res Commun. 1999. PMID: 10527867
-
Detection of reactive oxygen species derived from the family of NOX NADPH oxidases.Free Radic Biol Med. 2012 Nov 15;53(10):1903-18. doi: 10.1016/j.freeradbiomed.2012.09.002. Epub 2012 Sep 13. Free Radic Biol Med. 2012. PMID: 22982596 Review.
-
Electron spin resonance spectroscopy for the study of nanomaterial-mediated generation of reactive oxygen species.J Food Drug Anal. 2014 Mar;22(1):49-63. doi: 10.1016/j.jfda.2014.01.004. Epub 2014 Feb 1. J Food Drug Anal. 2014. PMID: 24673903 Free PMC article. Review.
Cited by
-
Activation of TRPC6 channels is essential for lung ischaemia-reperfusion induced oedema in mice.Nat Commun. 2012 Jan 31;3:649. doi: 10.1038/ncomms1660. Nat Commun. 2012. PMID: 22337127 Free PMC article.
-
Expression of NADPH oxidase (NOX) 5 in rabbit corneal stromal cells.PLoS One. 2012;7(4):e34440. doi: 10.1371/journal.pone.0034440. Epub 2012 Apr 12. PLoS One. 2012. PMID: 22511941 Free PMC article.
-
Oxygen-dependent regulation of ion channels: acute responses, post-translational modification, and response to chronic hypoxia.Pflugers Arch. 2021 Oct;473(10):1589-1602. doi: 10.1007/s00424-021-02590-7. Epub 2021 Jun 17. Pflugers Arch. 2021. PMID: 34142209 Review.
-
Mitochondrial Complex IV Subunit 4 Isoform 2 Is Essential for Acute Pulmonary Oxygen Sensing.Circ Res. 2017 Aug 4;121(4):424-438. doi: 10.1161/CIRCRESAHA.116.310482. Epub 2017 Jun 15. Circ Res. 2017. PMID: 28620066 Free PMC article.
-
Bypassing mitochondrial complex III using alternative oxidase inhibits acute pulmonary oxygen sensing.Sci Adv. 2020 Apr 15;6(16):eaba0694. doi: 10.1126/sciadv.aba0694. eCollection 2020 Apr. Sci Adv. 2020. PMID: 32426457 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
