

Review
doi: 10.1016/j.gde.2005.04.015.
Mechanisms underlying the transcriptional regulation of skeletal myogenesis
Affiliations
- PMID: 16055324
- PMCID: PMC1283108
- DOI: 10.1016/j.gde.2005.04.015
Item in Clipboard
Review
Mechanisms underlying the transcriptional regulation of skeletal myogenesis
Curr Opin Genet Dev.
2005 Oct.
Abstract
During skeletal myogenesis, chromatin-modifying enzymes are engaged at discrete genomic regions by transcription factors that recognize sequence-specific DNA motifs located at muscle gene regulatory regions. The composition of the chromatin-bound protein complexes and their temporally and spatially regulated recruitment influence gene expression. Recent findings are consistent with the concept that chromatin modifiers play an important role in regulating skeletal muscle gene expression and cellular differentiation.
Figures

The presence of specific protein complexes at the chromatin of muscle loci correlates with defined histone modifications and transcriptional activity. (a) In undifferentiated myoblasts, a protein complex containing MyoD, pCAF and Sir2 is detected at transcriptionally inactive muscle regulatory regions. Lysines 9 and 14 of histone H3 are hypoacetylated. Interaction of NAD+ with Sir2 is required to stimulate Sir2 deacetylase activity. (b) Upon induction of muscle differentiation, the protein complex is still detected on the chromatin, but lysines 9 and 14 become hyperacetylated and transcription is initiated. The [NAD+]/[NADH] ratio is reduced and nicotinamide (NAM) might inhibit Sir2. (c) The transcription factor YY1 recruits the Polycomb Ezh2 methyltransferase and HDAC1 deacetylase at specific inactive muscle loci. Lysine 27 of histone H3 is methylated by Ezh2, and several other lysines are hypoacetylated. The transcription factor Msx1 interacts with the linker histone H1 in a region where lysines 9 and 14 of histone H3 are hypoacetylated and lysine 9 is methylated. (d) Transcriptional activation is associated with loss of the YY1–Ezh2–HDAC1 complex and recruitment of the transcription factors SRF, MyoD and associated chromatin-remodeling SWI–SNF complex and histone acetyltransferases (HATs). The p38 kinase interacts with and phosphorylates the BAF60 subunit of SWI–SNF and phosphorylates E12/E47 and MEF2C, which, in turn, recruits the RNA polymerase II complex. Formation of the activating protein complex coincides with lysine hyperacetylation, lysine 27 demethylation and MyoD acetylation.
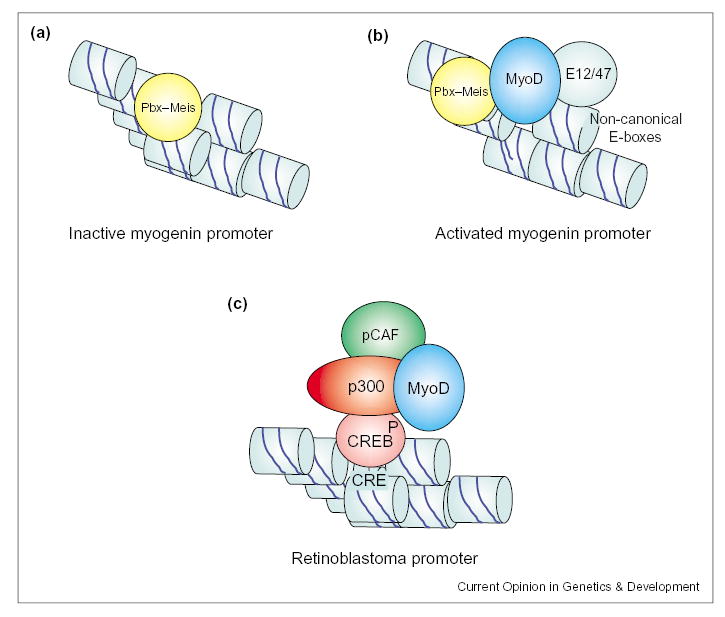
Indirect recruitment of MyoD at specific muscle promoters. (a) The myogenin promoter is constitutively occupied by a protein complex containing the homeodomain protein Pbx in undifferentiated myoblasts. (b) Interaction of MyoD–E12 heterodimers with non-canonical E-boxes of the myogenin promoter is stabilized through interaction with the Pbx complex, and coincides with transcriptional activation. (c) MyoD is indirectly recruited on the promoter of the retinoblastoma gene through interaction with a p300–pCAF complex associated with phosphorylated CREB bound to a DNA CRE element.
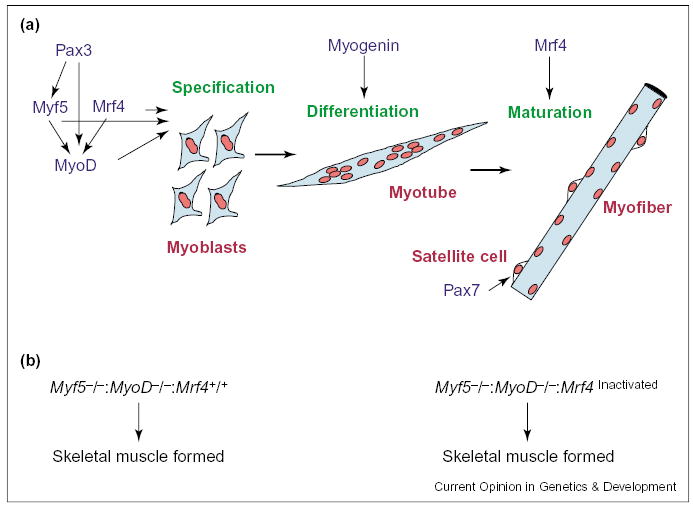
Hierarchical relationships of muscle regulatory factors. (a) Pax3, Myf5 and Mrf4 activate MyoD. Myf5, Mrf4 and MyoD can act as specification genes, whereas myogenin is required for differentiation of specified myoblasts into myotubes, and Mrf4 is involved in myofiber formation. Pax7 is required for satellite cell specification. (b) In the absence of both MyoD and Myf5, skeletal muscle is formed as long as expression of Mrf4 is not perturbed. Functional inactivation of Mrf4 — a consequence of genetic ablation of Myf5 in MyoD−/− Myf5−/− double mutant mice — results in the lack of skeletal muscle.
References
-
- Gill G, Something about SUMO inhibits transcription Curr Opin Genet Dev 2005, 15: in press. - PubMed
-
- Bitterman KJ, Anderson RM, Cohen HY, Latorre-Esteves M, Sinclair DA. Inhibition of silencing and accelerated aging by nicotinamide, a putative negative regulator of yeast sir2 and human SIRT1. J Biol Chem. 2002;277:45099–45107. - PubMed
-
- Fulco M, Schiltz RL, Iezzi S, King MT, Zhao P, Kashiwaya Y, Hoffman E, Veech RL, Sartorelli V. Sir2 regulates skeletal muscle differentiation as a potential sensor of the redox state. Mol Cell. 2003;12:51–62. - PubMed
-
- Thompson PR, Wang D, Wang L, Fulco M, Pediconi N, Zhang D, An W, Ge Q, Roeder RG, Wong J, et al. Regulation of the p300 HAT domain via a novel activation loop. Nat Struct Mol Biol. 2004;11:308–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
