

Surveillance of different recombination defects in mouse spermatocytes yields distinct responses despite elimination at an identical developmental stage
- PMID: 16055729
- PMCID: PMC1190256
- DOI: 10.1128/MCB.25.16.7203-7215.2005
Surveillance of different recombination defects in mouse spermatocytes yields distinct responses despite elimination at an identical developmental stage
Abstract
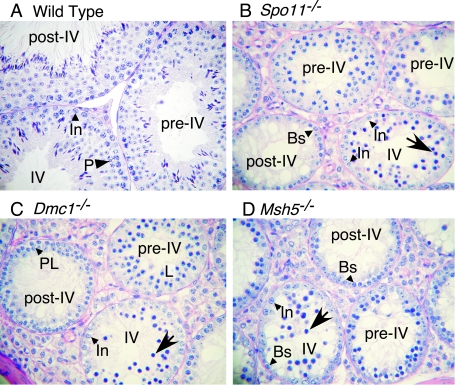
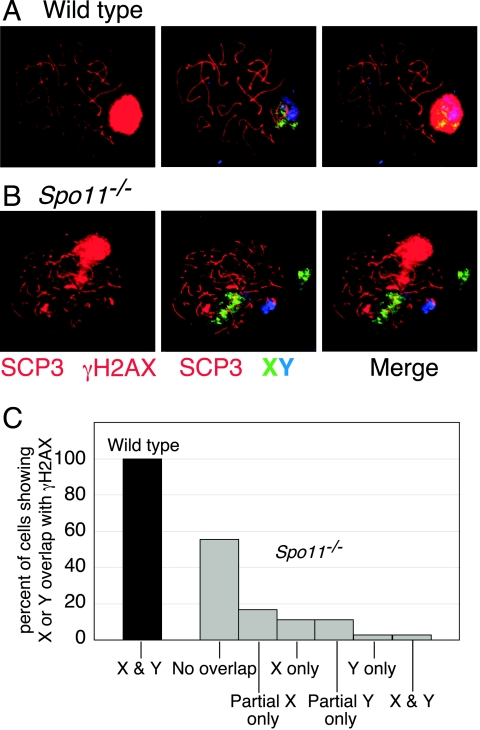
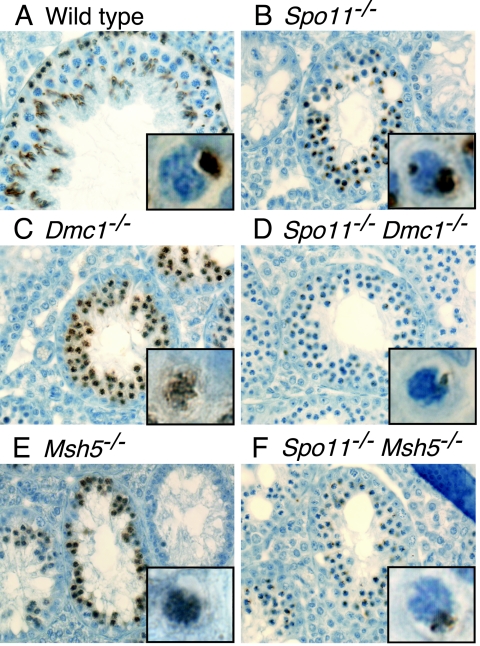
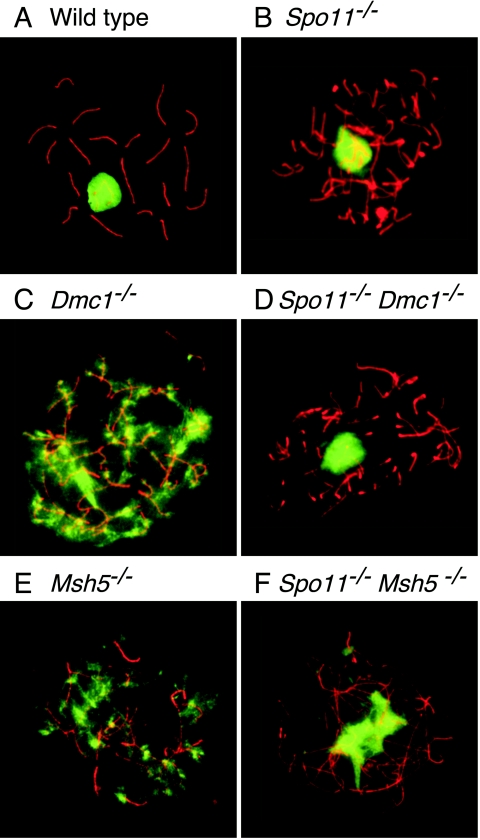
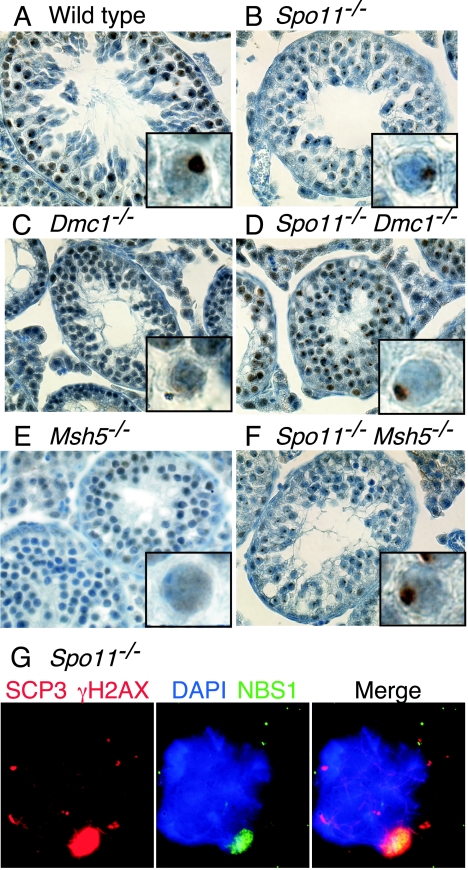
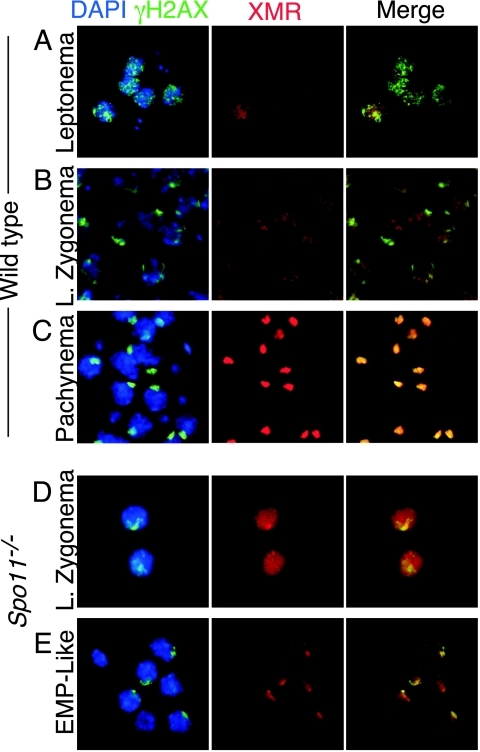
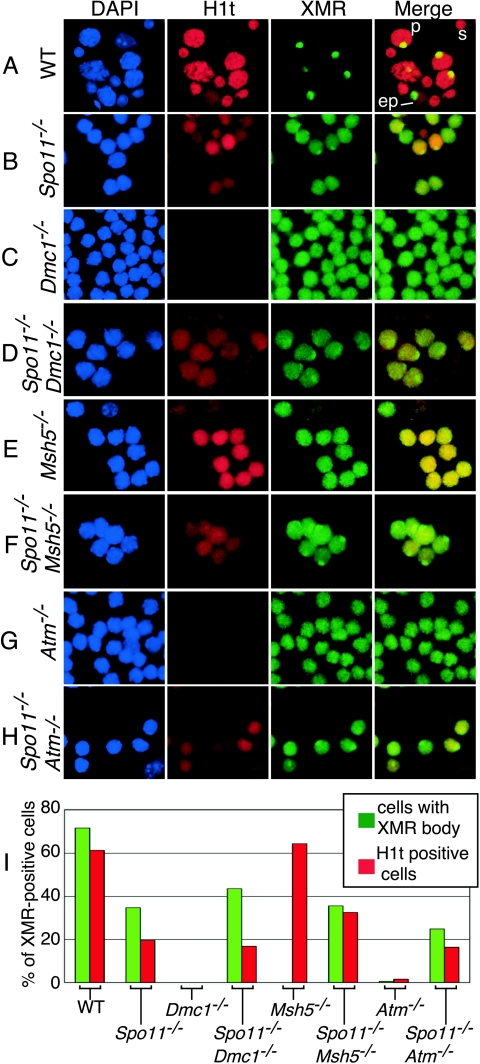
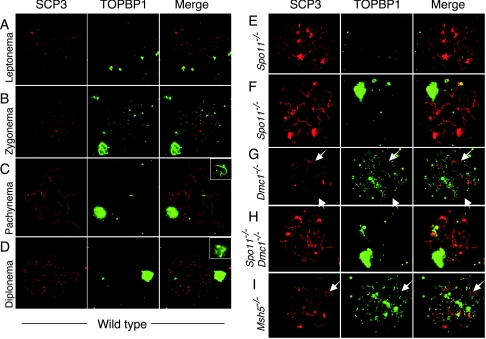
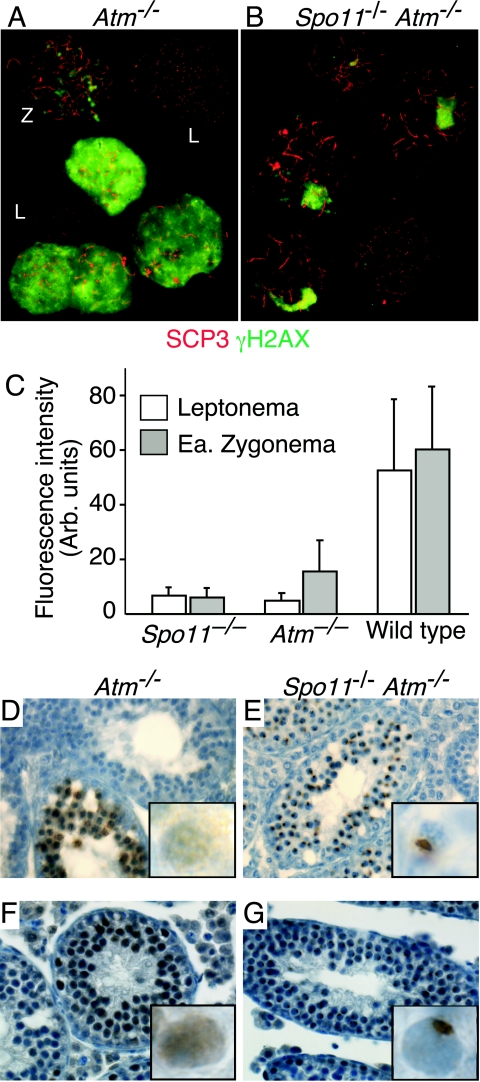
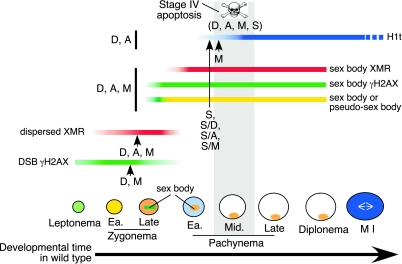
Fundamentally different recombination defects cause apoptosis of mouse spermatocytes at the same stage in development, stage IV of the seminiferous epithelium cycle, equivalent to mid-pachynema in normal males. To understand the cellular response(s) that triggers apoptosis, we examined markers of spermatocyte development in mice with different recombination defects. In Spo11(-)(/)(-) mutants, which lack the double-strand breaks (DSBs) that initiate recombination, spermatocytes express markers of early to mid-pachynema, forming chromatin domains that contain sex body-associated proteins but that rarely encompass the sex chromosomes. Dmc1(-)(/)(-) spermatocytes, impaired in DSB repair, appear to arrest at or about late zygonema. Epistasis analysis reveals that this earlier arrest is a response to unrepaired DSBs, and cytological analysis implicates the BRCT-containing checkpoint protein TOPBP1. Atm(-)(/)(-) spermatocytes show similarities to Dmc1(-)(/)(-) spermatocytes, suggesting that ATM promotes meiotic DSB repair. Msh5(-)(/)(-) mutants display a set of characteristics distinct from these other mutants. Thus, despite equivalent stages of spermatocyte elimination, different recombination-defective mutants manifest distinct responses, providing insight into surveillance mechanisms in male meiosis.
Figures
References
-
- Ashley, T., C. Westphal, A. Plug-de Maggio, and D. G. de Rooij. 2004. The mammalian mid-pachytene checkpoint: meiotic arrest in spermatocytes with a mutation in Atm alone or in combination with a Trp53 (p53) or Cdkn1a (p21/cip1) mutation. Cytogenet. Genome Res. 107:256-262. - PubMed
-
- Bakkenist, C. J., and M. B. Kastan. 2003. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421:499-506. - PubMed
-
- Barlow, C., M. Liyanage, P. B. Moens, M. Tarsounas, K. Nagashima, K. Brown, S. Rottinghaus, S. P. Jackson, D. Tagle, T. Ried, and A. Wynshaw-Boris. 1998. Atm deficiency results in severe meiotic disruption as early as leptonema of prophase I. Development 125:4007-4017. - PubMed
-
- Baudat, F., K. Manova, J. P. Yuen, M. Jasin, and S. Keeney. 2000. Chromosome synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol. Cell 6:989-998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous