

Neurexophilin 3 is highly localized in cortical and cerebellar regions and is functionally important for sensorimotor gating and motor coordination
- PMID: 16055736
- PMCID: PMC1190246
- DOI: 10.1128/MCB.25.16.7278-7288.2005
Neurexophilin 3 is highly localized in cortical and cerebellar regions and is functionally important for sensorimotor gating and motor coordination
Abstract
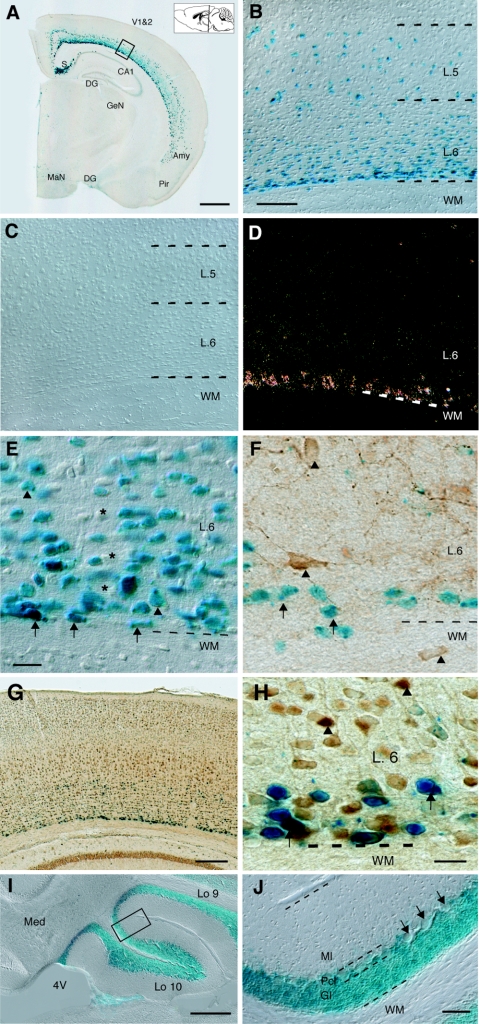
Neurexophilin 3 (Nxph3) is a specific ligand of synaptic alpha-neurexins that are essential for efficient neurotransmitter release. Previous biochemical work demonstrated that Nxph3 interacts with an extracellular domain of alpha-neurexins in a tight complex; however, no information is available on the localization or functional role of Nxph3 in the brain. Here, we generated lacZ reporter gene knock-in mice to investigate the distribution of Nxph3 at the single-cell level and Nxph3 knockout mice to examine its functional importance. Nxph3 expression was restricted mostly to subplate-derived neurons in cortical layer 6b, granule cells in the vestibulocerebellum, and Cajal-Retzius cells during development. Colabeling experiments demonstrated that neurons expressing Nxph3 do not belong to a uniform cell type. Morphological analyses and systematic behavioral testing of knockout mice revealed no anatomical defects but uncovered remarkable functional abnormalities in sensory information processing and motor coordination, evident by increased startle response, reduced prepulse inhibition, and poor rotarod performance. Since Nxph3-deficient mice behaved normally while performing a number of other tasks, our data suggest an important role for Nxph3 as a locally and temporally regulated neuropeptide-like molecule, presumably acting in a complex with alpha-neurexins in select neuronal circuits.
Figures






References
-
- Arber, S. 2004. Subplate neurons: bridging the gap to function in the cortex. Trends Neurosci. 27:111-113. - PubMed
-
- Carlson, S., and J. F. Willott. 1998. Caudal pontine reticular formation of C57BL/6J mice: responses to startle stimuli, inhibition by tones, and plasticity. J. Neurophysiol. 79:2603-2614. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases