

Essential role of CIB1 in regulating PAK1 activation and cell migration
- PMID: 16061695
- PMCID: PMC2171473
- DOI: 10.1083/jcb.200502090
Essential role of CIB1 in regulating PAK1 activation and cell migration
Abstract
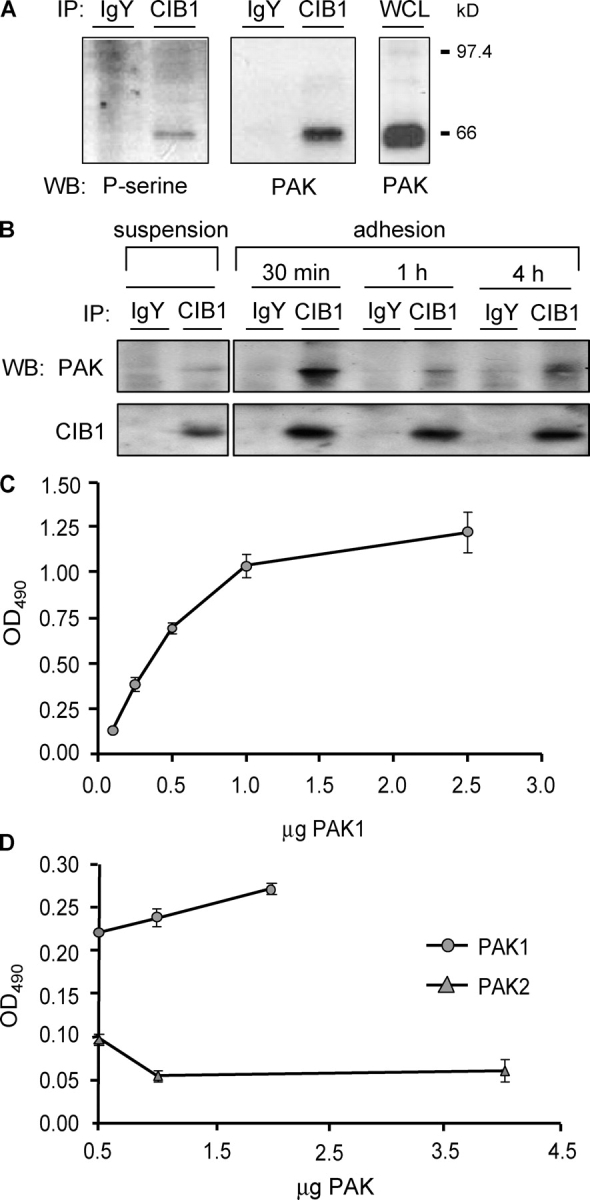
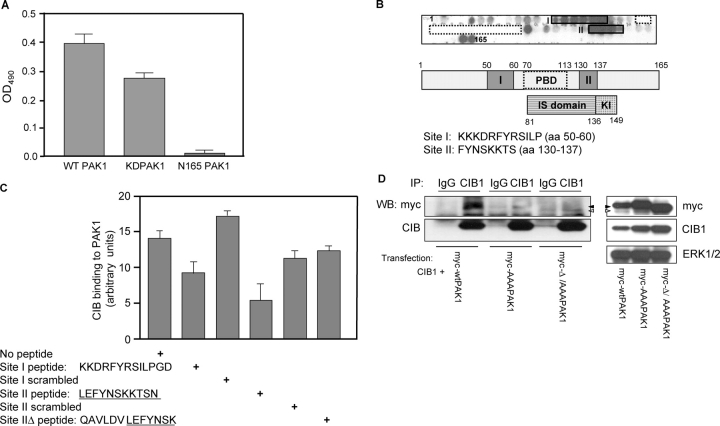
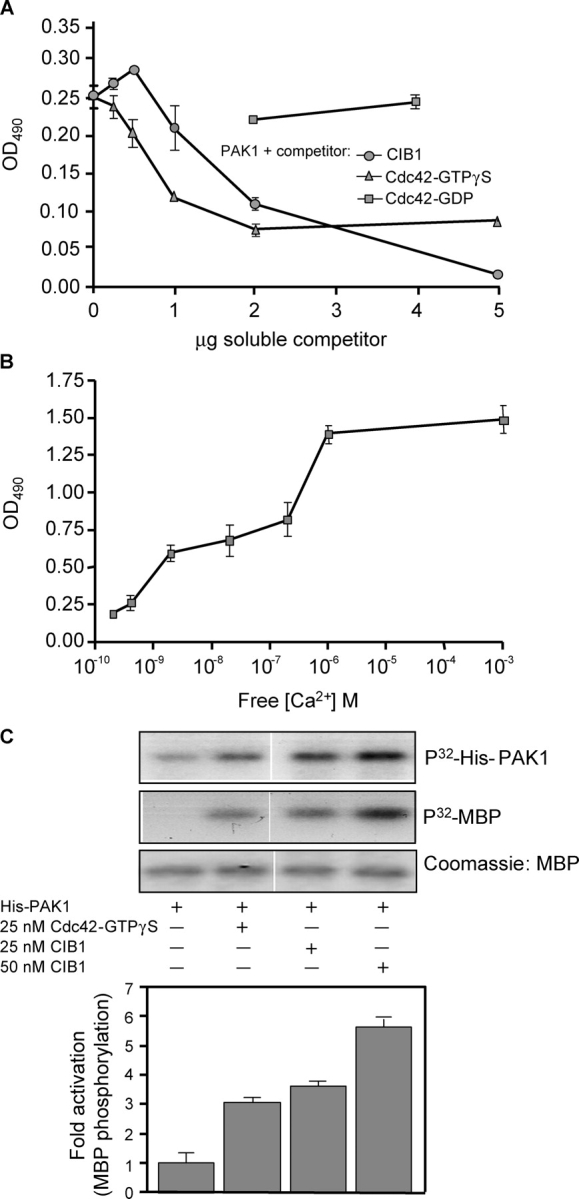
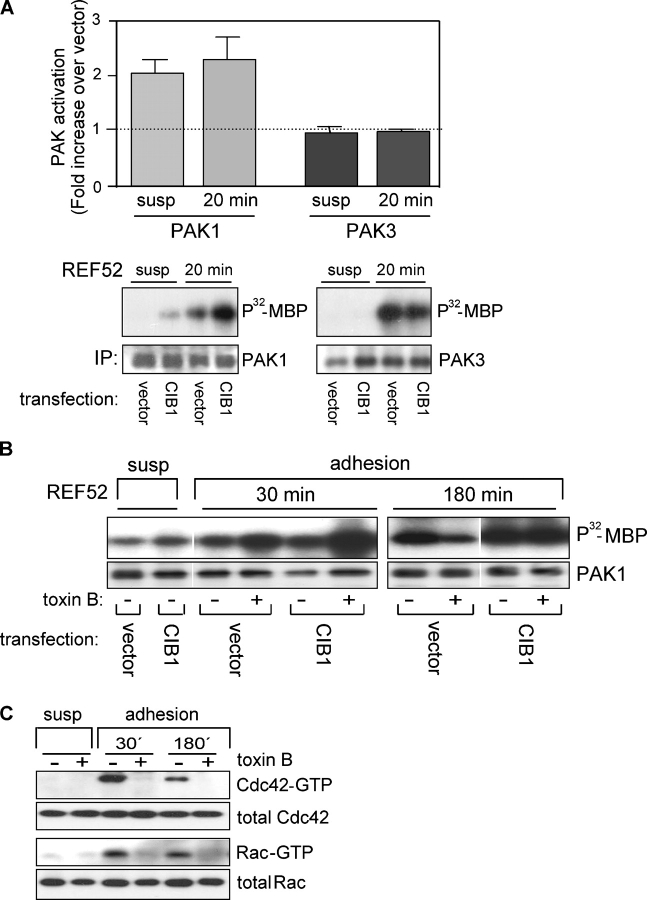
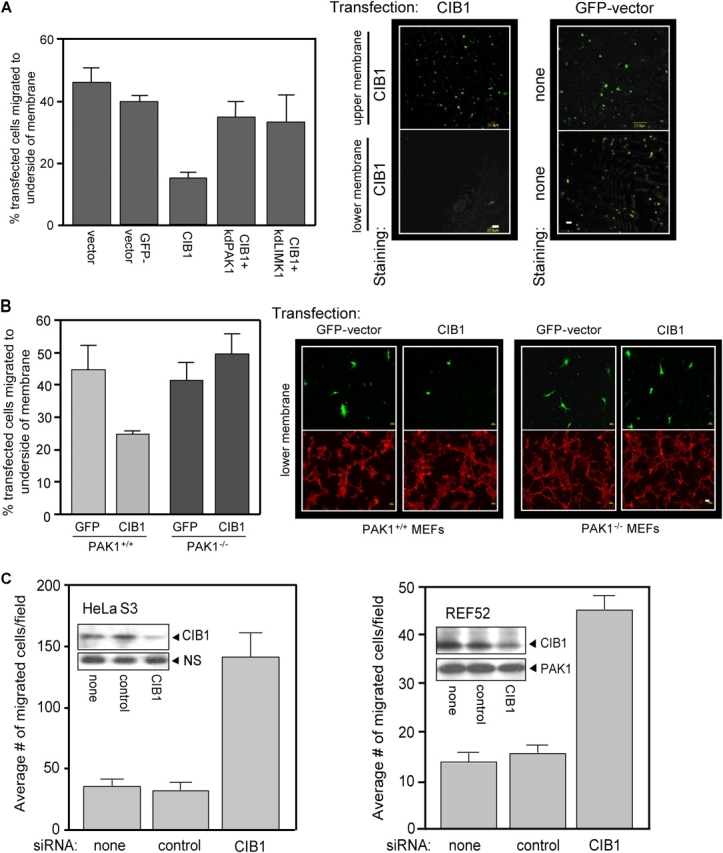
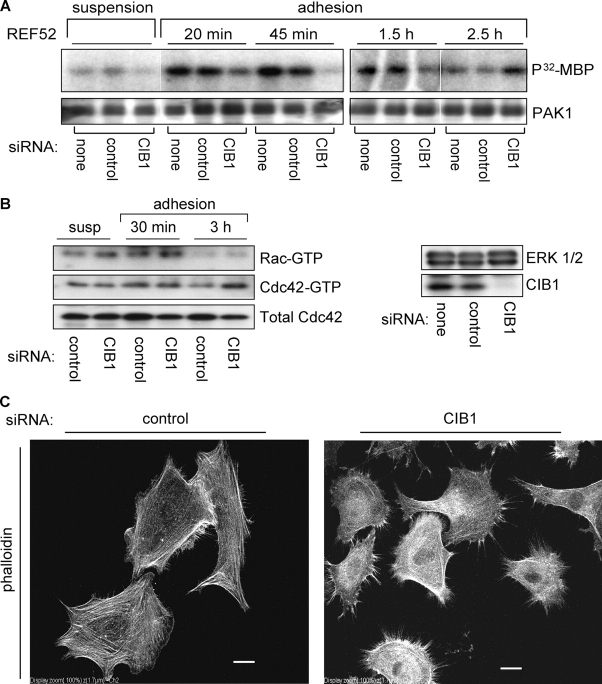
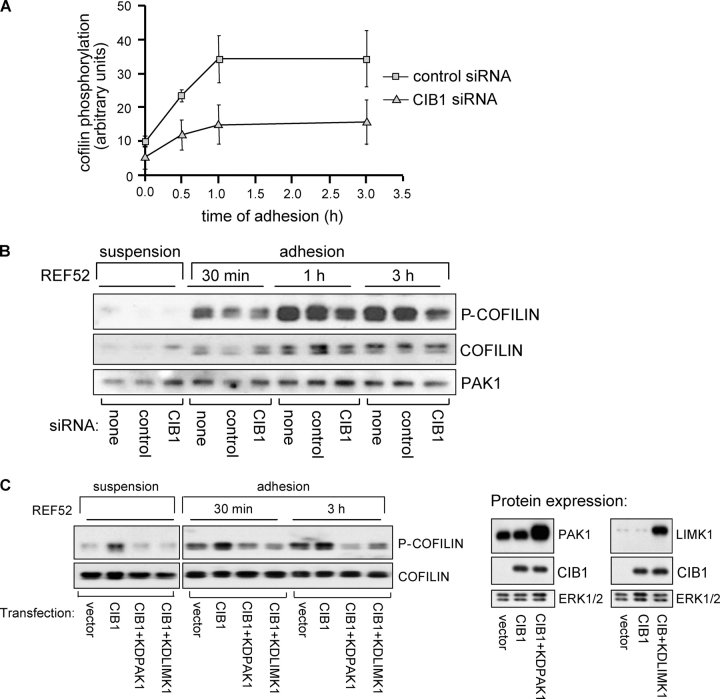
p21-activated kinases (PAKs) regulate many cellular processes, including cytoskeletal rearrangement and cell migration. In this study, we report a direct and specific interaction of PAK1 with a 22-kD Ca2+-binding protein, CIB1, which results in PAK1 activation both in vitro and in vivo. CIB1 binds to PAK1 within discrete regions surrounding the inhibitory switch domain in a calcium-dependent manner, providing a potential mechanism of CIB1-induced PAK1 activation. CIB1 overexpression significantly decreases cell migration on fibronectin as a result of a PAK1-and LIM kinase-dependent increase in cofilin phosphorylation. Conversely, the RNA interference-mediated depletion of CIB1 increases cell migration and reduces normal adhesion-induced PAK1 activation and cofilin phosphorylation. Together, these results demonstrate that endogenous CIB1 is required for regulated adhesion-induced PAK1 activation and preferentially induces a PAK1-dependent pathway that can negatively regulate cell migration. These results point to CIB1 as a key regulator of PAK1 activation and signaling.
Figures
References
-
- Adam, L., R. Vadlamudi, M. Mandal, J. Chernoff, and R. Kumar. 2000. Regulation of microfilament reorganization and invasiveness of breast cancer cells by kinase dead p21-activated kinase-1. J. Biol. Chem. 275:12041–12050. - PubMed
-
- Arber, S., F.A. Barbayannis, H. Hanser, C. Schneider, C.A. Stanyon, O. Bernard, and P. Caroni. 1998. Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature. 393:805–809. - PubMed
-
- Bagheri-Yarmand, R., M. Mandal, A.H. Taludker, R.A. Wang, R.K. Vadlamudi, H.J. Kung, and R. Kumar. 2001. Etk/Bmx tyrosine kinase activates Pak1 and regulates tumorigenicity of breast cancer cells. J. Biol. Chem. 276:29403–29409. - PubMed
-
- Bagrodia, S., S.J. Taylor, C.L. Creasy, J. Chernoff, and R.A. Cerione. 1995. Identification of a mouse p21Cdc42/Rac activated kinase. J. Biol. Chem. 270:22731–22737. - PubMed
-
- Bagrodia, S., S.J. Taylor, K.A. Jordon, L. Van Aelst, and R.A. Cerione. 1998. A novel regulator of p21-activated kinases. J. Biol. Chem. 273:23633–23636. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
