

The epidermal barrier function is dependent on the serine protease CAP1/Prss8
- PMID: 16061697
- PMCID: PMC2171460
- DOI: 10.1083/jcb.200501038
The epidermal barrier function is dependent on the serine protease CAP1/Prss8
Abstract
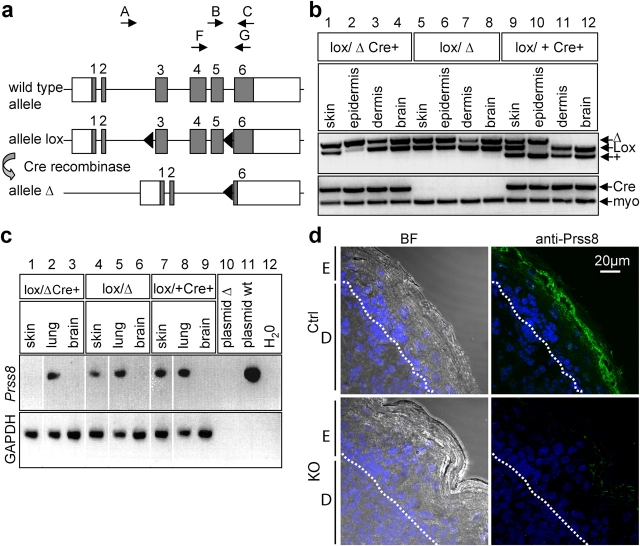
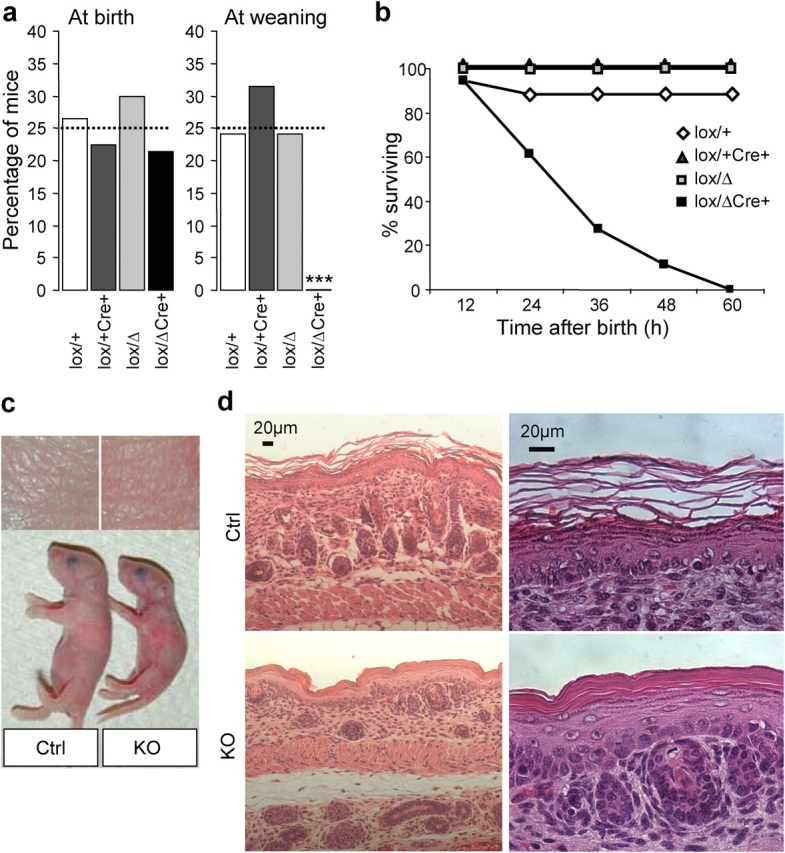
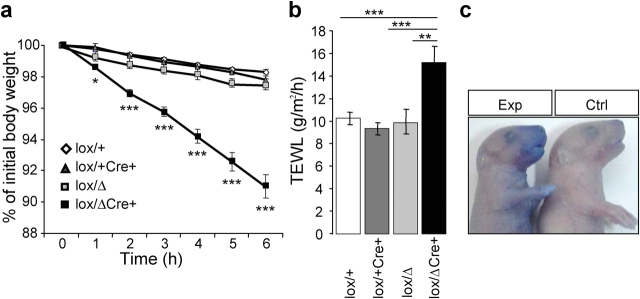
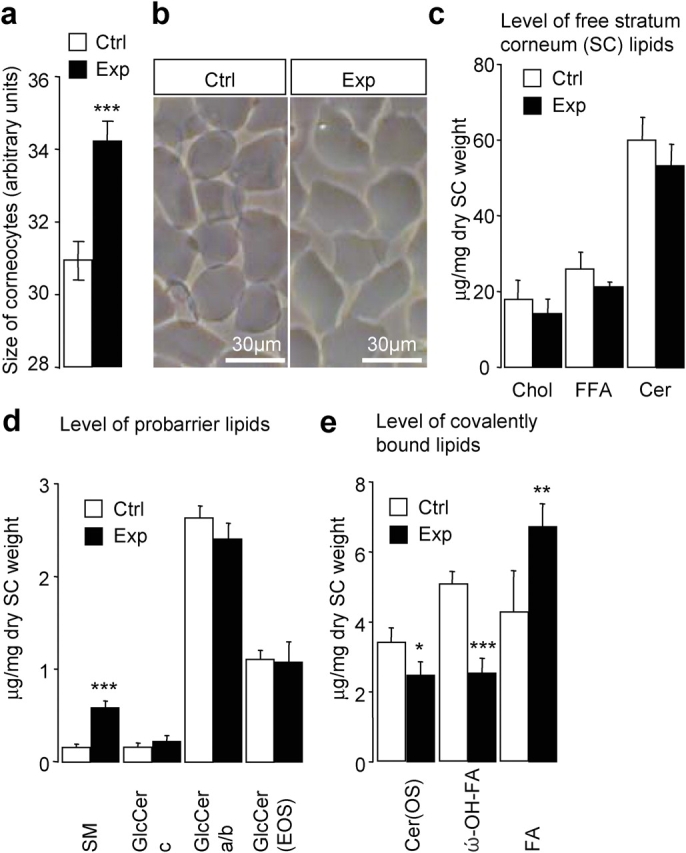
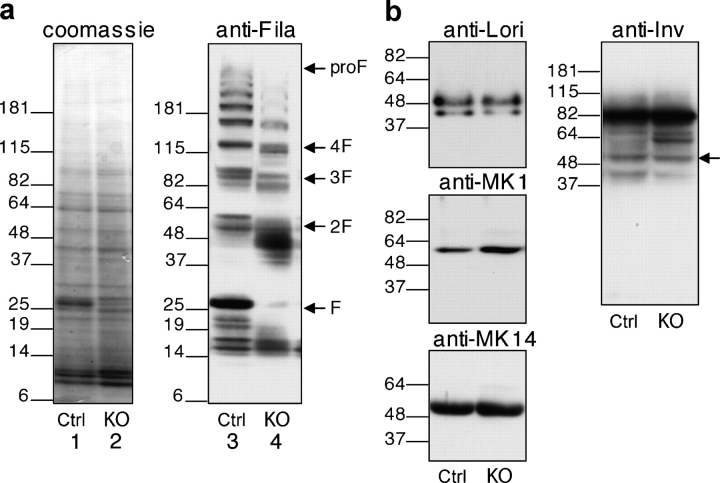
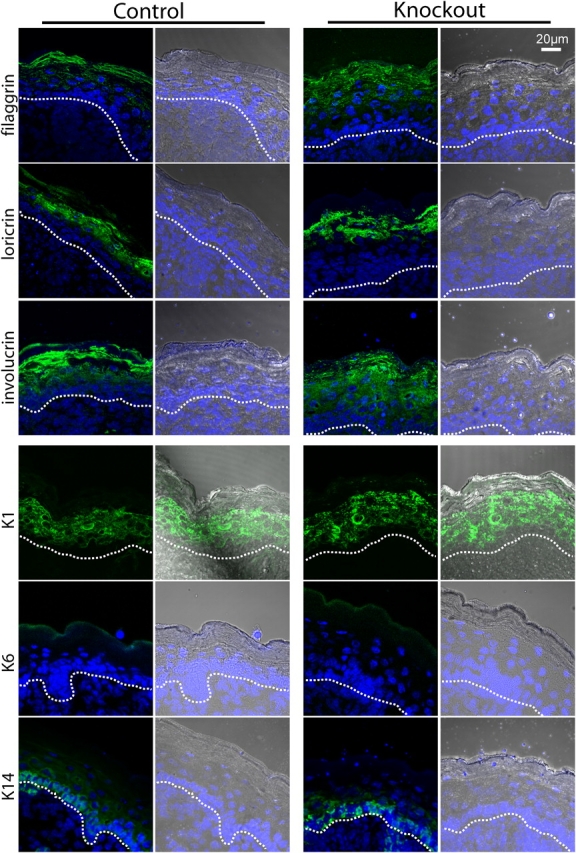
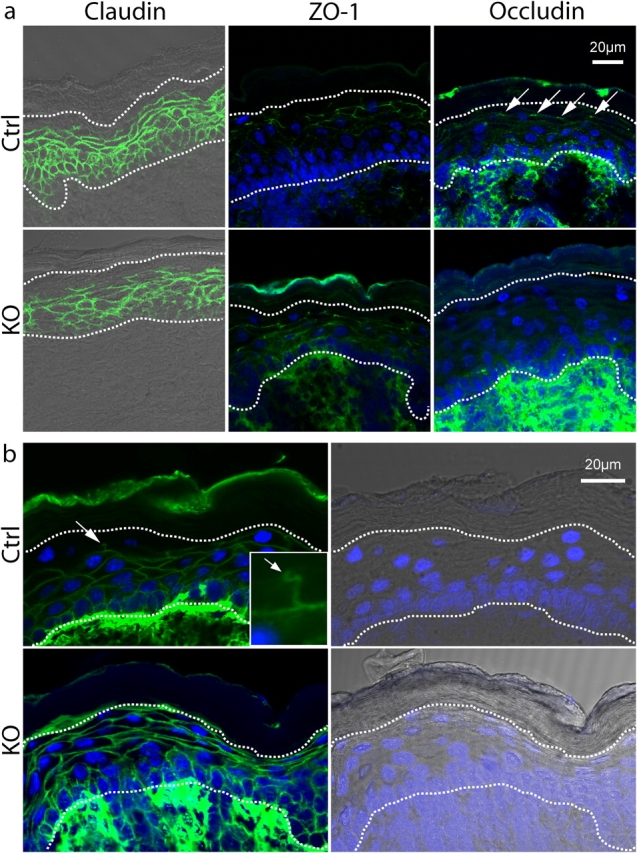
Serine proteases are proteolytic enzymes that are involved in the regulation of various physiological processes. We generated mice lacking the membrane-anchored channel-activating serine protease (CAP) 1 (also termed protease serine S1 family member 8 [Prss8] and prostasin) in skin, and these mice died within 60 h after birth. They presented a lower body weight and exhibited severe malformation of the stratum corneum (SC). This aberrant skin development was accompanied by an impaired skin barrier function, as evidenced by dehydration and skin permeability assay and transepidermal water loss measurements leading to rapid, fatal dehydration. Analysis of differentiation markers revealed no major alterations in CAP1/Prss8-deficient skin even though the epidermal deficiency of CAP1/Prss8 expression disturbs SC lipid composition, corneocyte morphogenesis, and the processing of profilaggrin. The examination of tight junction proteins revealed an absence of occludin, which did not prevent the diffusion of subcutaneously injected tracer (approximately 600 D) toward the skin surface. This study shows that CAP1/Prss8 expression in the epidermis is crucial for the epidermal permeability barrier and is, thereby, indispensable for postnatal survival.
Figures
References
-
- Barel, A.O., and P. Clarys. 1995. Study of the stratum corneum barrier function by transepidermal water loss measurements: comparison between two commercial instruments: Evaporimeter and Tewameter. Skin Pharmacol. 8:186–195. - PubMed
-
- Brandner, J.M., S. Kief, C. Grund, M. Rendl, P. Houdek, C. Kuhn, E. Tschachler, W.W. Franke, and I. Moll. 2002. Organization and formation of the tight junction system in human epidermis and cultured keratinocytes. Eur. J. Cell Biol. 81:253–263. - PubMed
-
- Brouard, M., M. Casado, S. Djelidi, Y. Barrandon, and N. Farman. 1999. Epithelial sodium channel in human epidermal keratinocytes: expression of its subunits and relation to sodium transport and differentiation. J. Cell Sci. 112:3343–3352. - PubMed
-
- Chen, L.M., X. Zhang, and K.X. Chai. 2004. Regulation of prostasin expression and function in the prostate. Prostate. 59:1–12. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
