

Selective targeting of regulatory T cells with CD28 superagonists allows effective therapy of experimental autoimmune encephalomyelitis
- PMID: 16061730
- PMCID: PMC2213080
- DOI: 10.1084/jem.20051060
Selective targeting of regulatory T cells with CD28 superagonists allows effective therapy of experimental autoimmune encephalomyelitis
Abstract
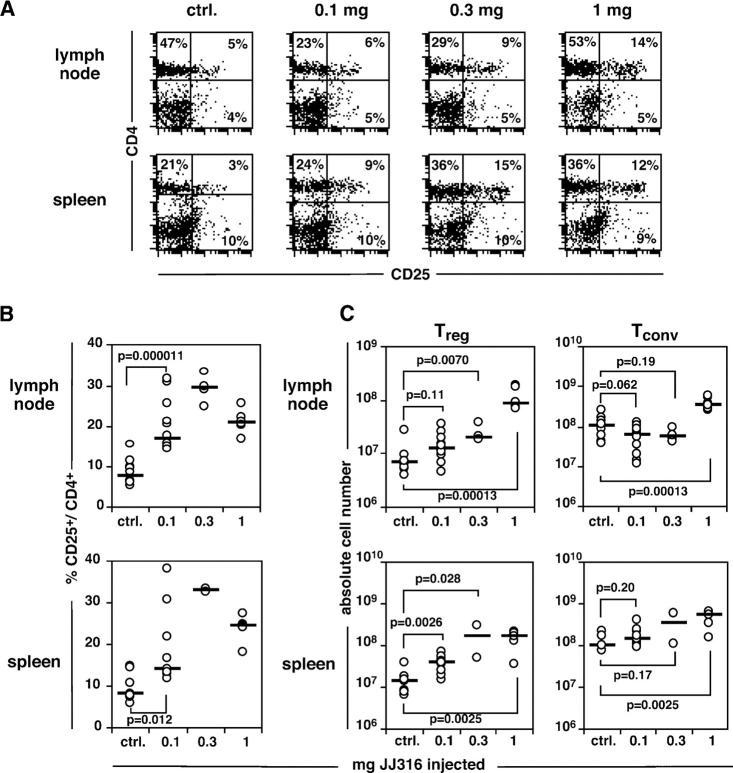
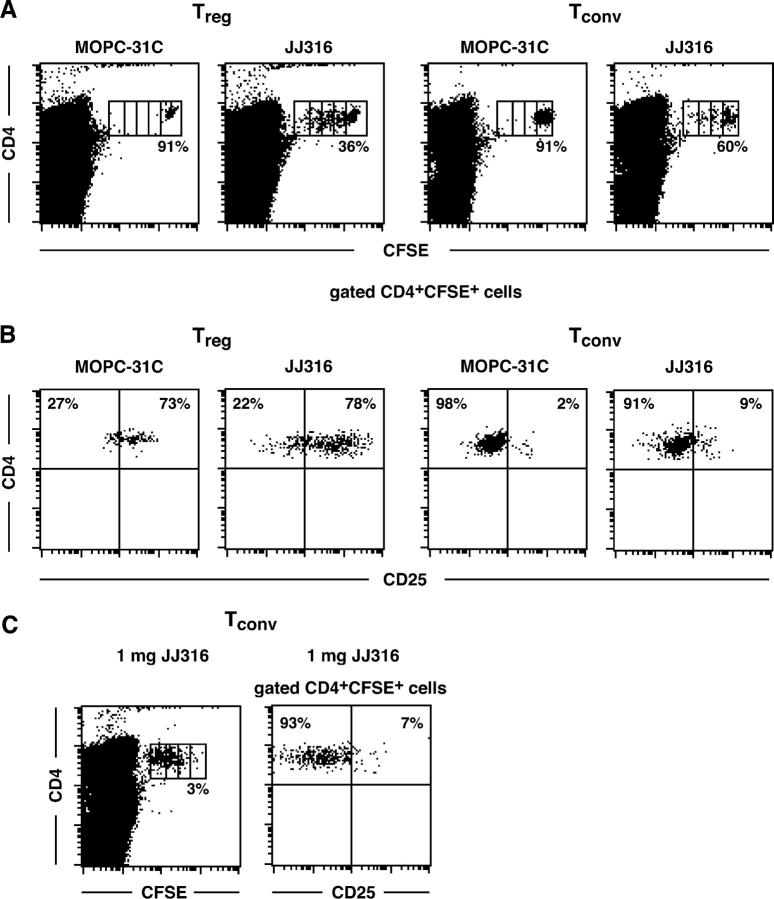
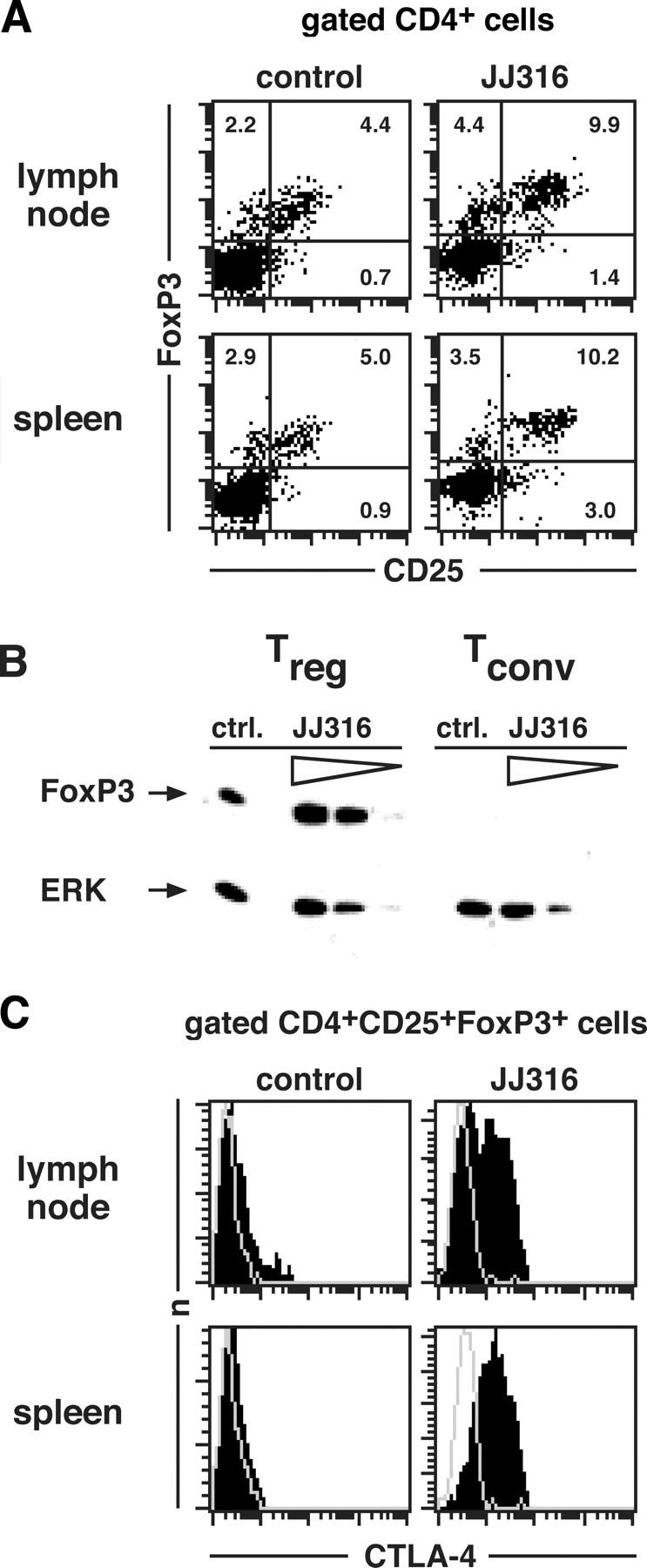
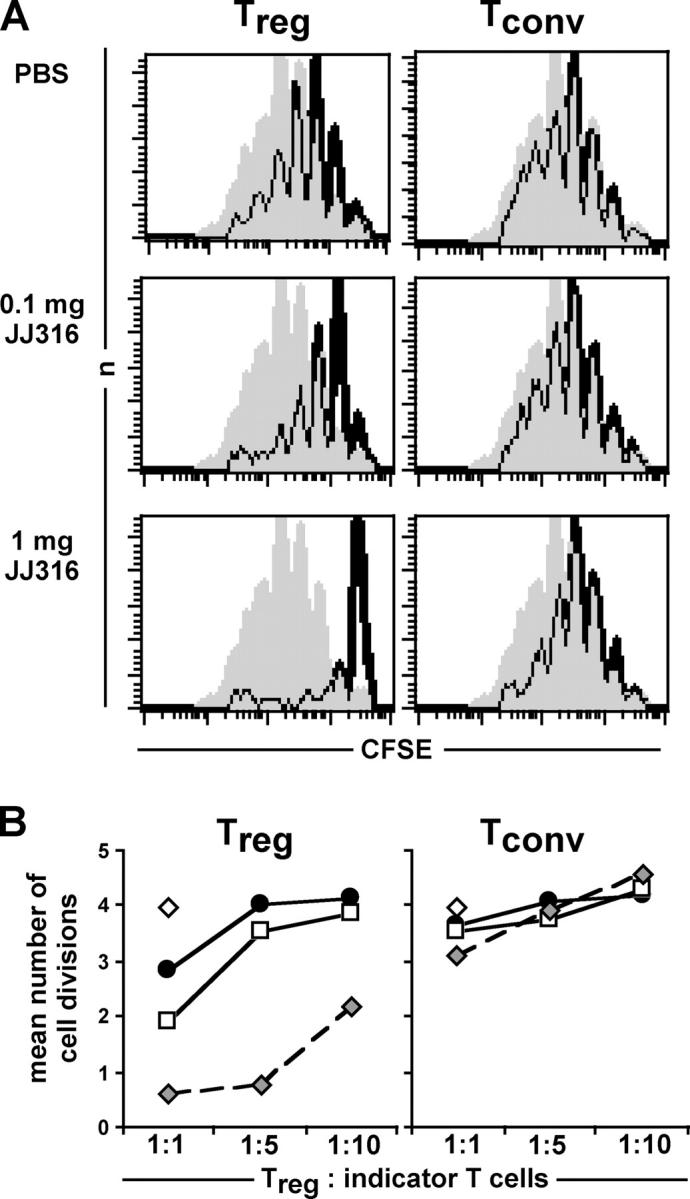
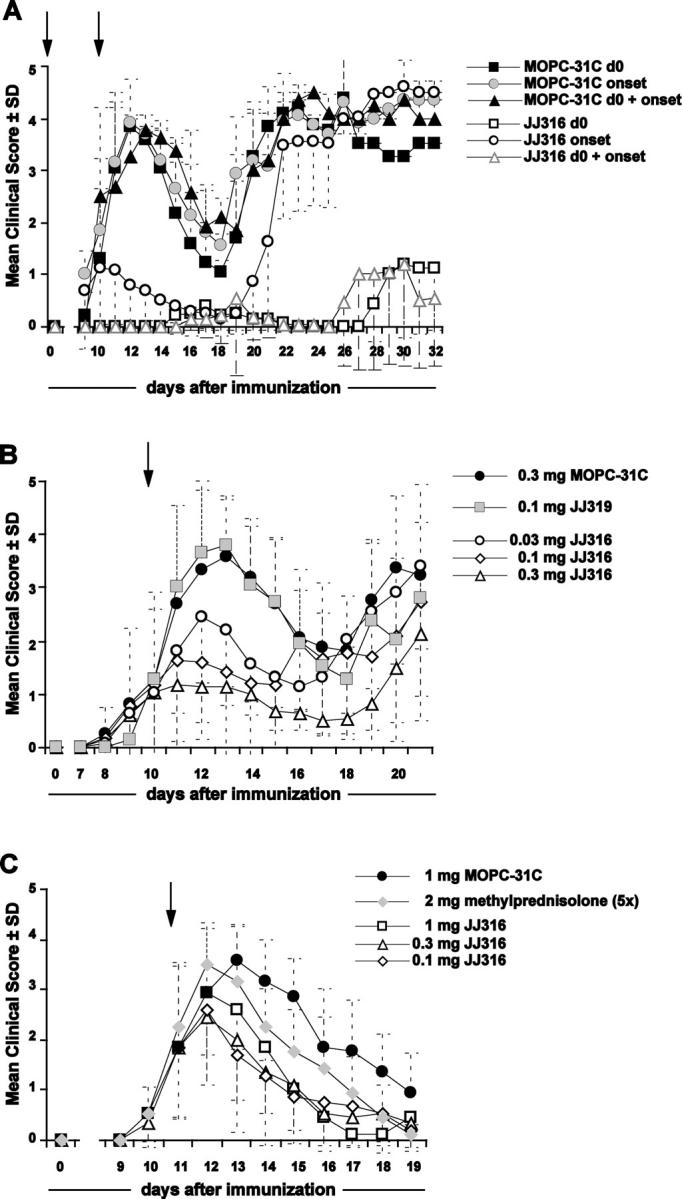
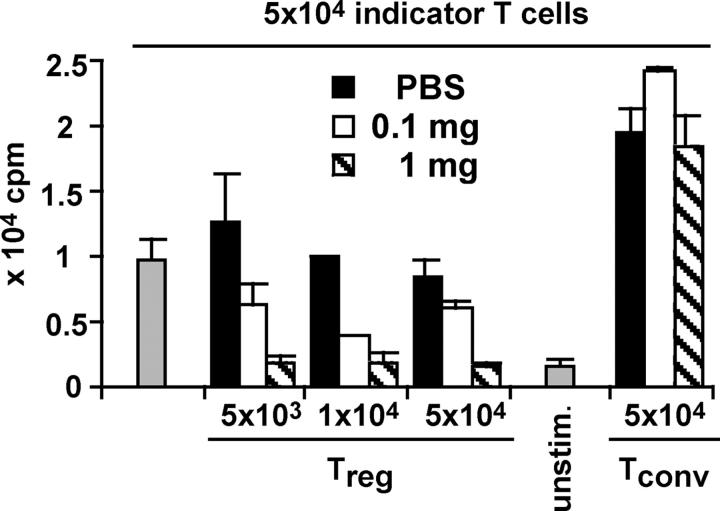
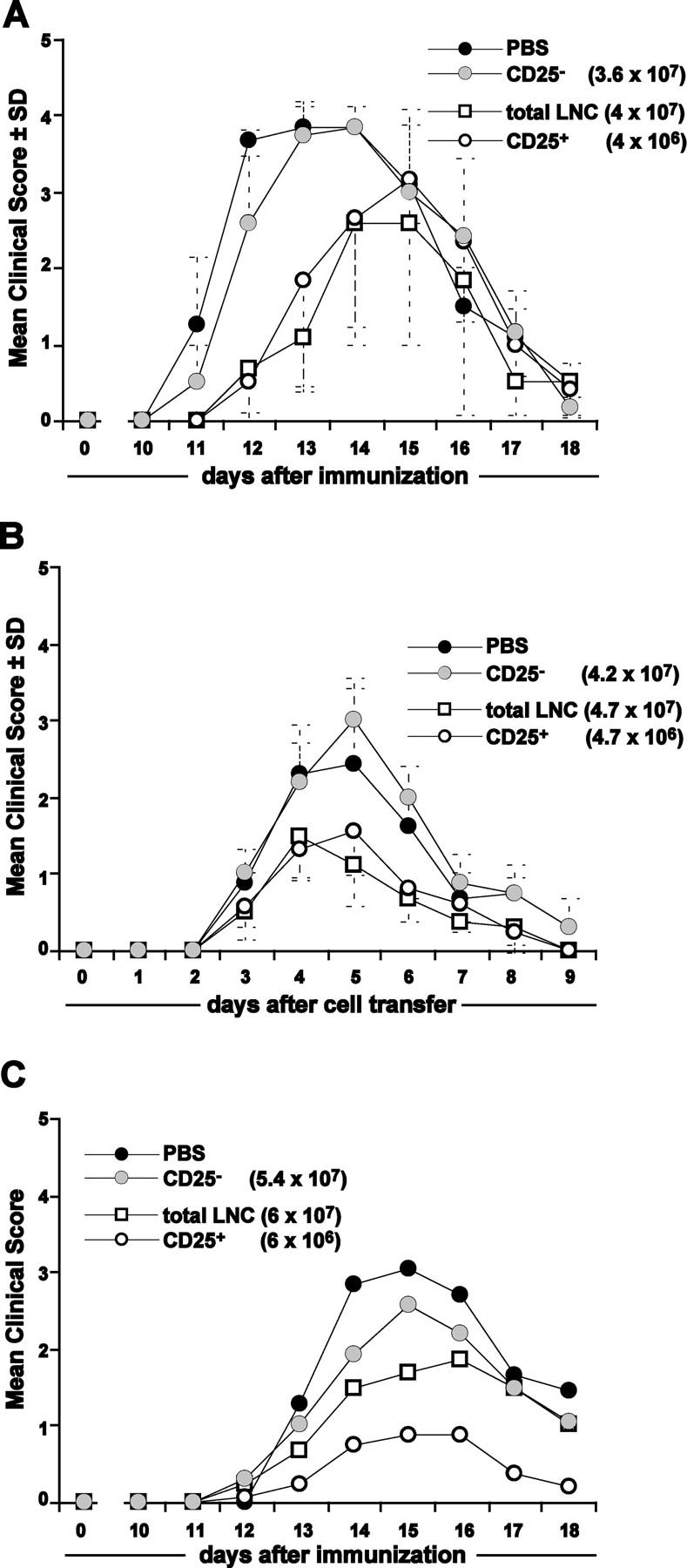
CD4+CD25+ regulatory T cells (T reg cells) play a key role in controlling autoimmunity and inflammation. Therefore, therapeutic agents that are capable of elevating numbers or increasing effector functions of this T cell subset are highly desirable. In a previous report we showed that a superagonistic monoclonal antibody specific for rat CD28 (JJ316) expands and activates T reg cells in vivo and upon short-term in vitro culture. Here we demonstrate that application of very low dosages of the CD28 superagonist into normal Lewis rats is sufficient to induce T reg cell expansion in vivo without the generalized lymphocytosis observed with high dosages of JJ316. Single i.v. administration of a low dose of the CD28 superagonist into Dark Agouti (DA) rats or Lewis rats that suffered from experimental autoimmune encephalomyelitis (EAE) proved to be highly and equally efficacious as high-dose treatment. Finally, we show that T reg cells that were isolated from CD28-treated animals displayed enhanced suppressive activity toward myelin basic protein-specific T cells in vitro, and, upon adoptive transfer, protected recipients from EAE. Our data indicate that this class of CD28-specific monoclonal antibodies targets CD4+CD25+ T reg cells and provides a novel means for the effective treatment of multiple sclerosis and other autoimmune diseases.
Figures
References
-
- Walker, L.S., and A.K. Abbas. 2002. The enemy within: keeping self-reactive T cells at bay in the periphery. Nat. Rev. Immunol. 2:11–19. - PubMed
-
- Sakaguchi, S., N. Sakaguchi, M. Asano, M. Itoh, and M. Toda. 1995. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 155:1151–1164. - PubMed
-
- Suri-Payer, E., A.Z. Amar, A.M. Thornton, and E.M. Shevach. 1998. CD4+CD25+ T cells inhibit both the induction and effector function of autoreactive T cells and represent a unique lineage of immunoregulatory cells. J. Immunol. 160:1212–1218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
