

Review
doi: 10.1172/JCI25495.
ATP-sensitive potassium channelopathies: focus on insulin secretion
Affiliations
- PMID: 16075046
- PMCID: PMC1180549
- DOI: 10.1172/JCI25495
Item in Clipboard
Review
ATP-sensitive potassium channelopathies: focus on insulin secretion
J Clin Invest.
2005 Aug.
Abstract
ATP-sensitive potassium (K(ATP)) channels, so named because they are inhibited by intracellular (ATP), play key physiological roles in many tissues. In pancreatic beta cells, these channels regulate glucose-dependent insulin secretion and serve as the target for sulfonylurea drugs used to treat type 2 diabetes. This review focuses on insulin secretory disorders, such as congenital hyperinsulinemia and neonatal diabetes, that result from mutations in K(ATP) channel genes. It also considers the extent to which defective regulation of K(ATP) channel activity contributes to the etiology of type 2 diabetes.
Figures
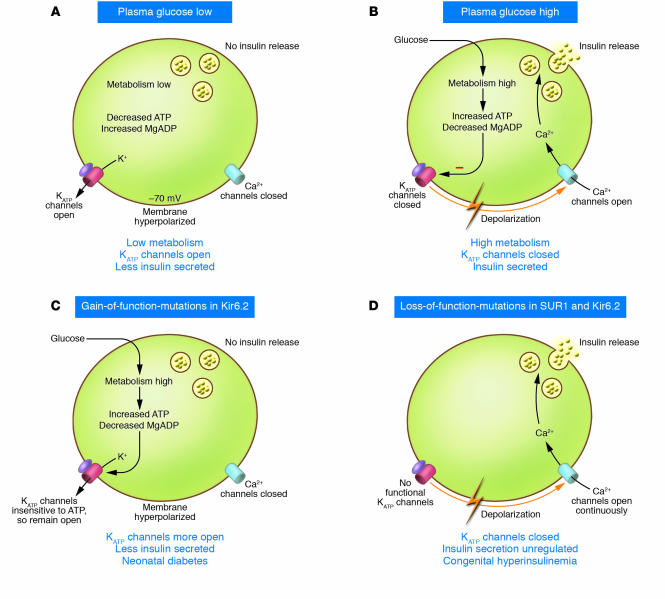
The KATP channel couples glucose metabolism to insulin secretion. Glucose enters the cell via the GLUT2 transporter, and via glycolytic and mitochondrial metabolism leads to an increase in ATP and a fall in MgADP in the immediate vicinity of the KATP channel. This results in KATP channel closure, membrane depolarization, opening of voltage-gated Ca2+ channels, Ca2+ influx, and exocytosis of insulin granules.
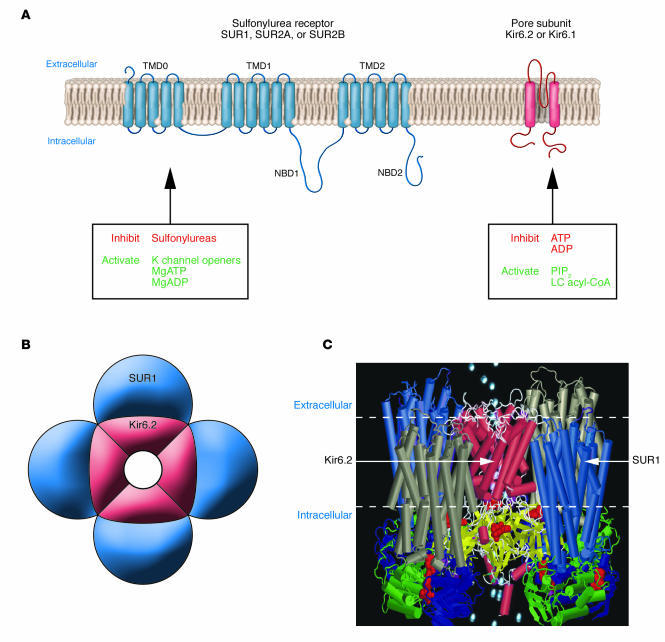
Molecular structure of the KATP channel. (A) Schematic representation of the transmembrane topology of a single SURx (left) or Kir6.x (right) subunit. Mg-nucleotide binding/hydrolysis at the nucleotide-binding domains (NBD1, NBD2) of SUR stimulates channel activity. Sulfonylureas (stimulatory) and K channel openers (inhibitory) also bind to SUR1. Binding of ATP or ADP to Kir6.2 closes the pore, an effect that does not require Mg2+. Conversely, binding of phospholipids such as PIP2, or long-chain (LC) acyl-coAs, stimulates KATP channel activity and decreases its ATP sensitivity. (B) Schematic representation of the octameric KATP channel complex viewed in cross section. Four Kir6.2 subunits come together to form the pore through which K+ ions move, and each is associated with a regulatory SURx subunit. (C) Model of how SUR1 and Kir6.2 might assemble to form the KATP channel. The SUR model is described in ref. and the Kir6.2 model in ref. . The model illustrates that the channel complex contains 4 ATP-binding sites (on Kir6.2) and 8 Mg nucleotide–binding sites (on SUR1).
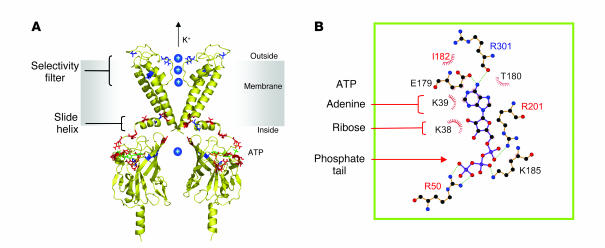
Location of disease-causing mutations in Kir6.2. (A) Structural model of Kir6.2 (116) viewed from the side. For clarity, only 2 transmembrane domains, and 2 separate cytosolic domains, are shown. Residues mutated in neonatal diabetes are shown in red, and those that cause hyperinsulinism of infancy in blue. ATP (green) is docked into its binding site. Of the residues implicated in neonatal diabetes, R50, R201, Y330C, and F333I lie close to the ATP-binding site; F35, C42, and E332K at the interface between Kir6.2 subunits; Q52 and G53 in a region postulated to interface with SUR1; and V59, C166, and I296L within regions of the channel involved in gating. (B) Close-up of the putative ATP-binding site with residues lying within 3.5 Å of ATP indicated. Residues mutated in neonatal diabetes are shown in red. Part B is adapted with permission from The EMBO Journal (116).
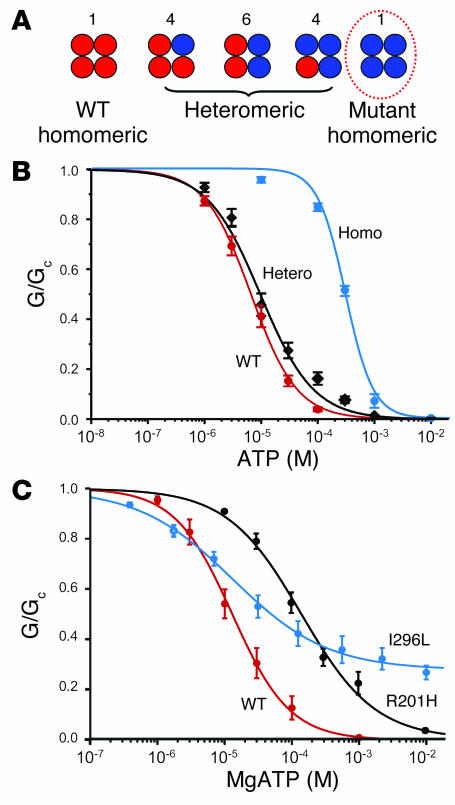
PNDM mutations reduce channel inhibition by ATP. (A) Schematic of the mixture of channels with different subunit compositions expected when WT and mutant Kir6.2 are coexpressed (as in the heterozygous state). The relative numbers of the channel types expected if WT and mutant subunits segregate independently (i.e., follow a binomial distribution) are indicated above the figure. The circle indicates the only channel type predicted to show a substantial change in ATP sensitivity if the mutation affects ATP binding (see text). (B) Mean relationship between [ATP] and KATP current (G), expressed relative to the conductance in the absence of nucleotide (GC) for Kir6.2/SUR1 (red, n = 6) and heterozygous (black, n = 6) and homomeric (blue, n = 6) Kir6.2-R201H/SUR1 channels. The smooth curves are the best fit to the Hill equation. The IC50 was 7 μM, 12 μM, and 300 μM for WT, heterozygous R201H, and homomeric R201H channels, respectively. Data were obtained in the absence of Mg2+. Part B reproduced with permission from Proceedings of the National Academy of Sciences of the United States of America (83). (C) Mean relationship between [MgATP] and KATP current, expressed relative to the conductance in the absence of nucleotide for Kir6.2/SUR1 (red, n = 6) and heterozygous Kir6.2-R201H (black, n = 5) and heterozygous Kir6.2-I296L/SUR1 (blue, n = 5) channels. The smooth curves are the best fit to the Hill equation. The IC50 was 13 μM, 140 μM, and 50 μM for WT, heterozygous R201H, and heterozygous I296L channels, respectively. Data in C are from refs. and .
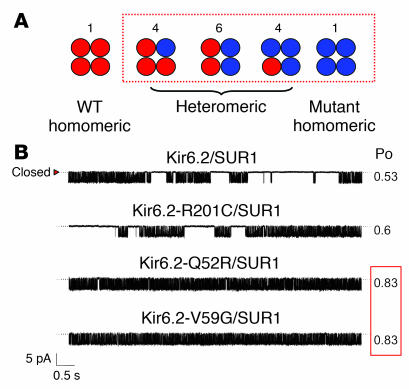
Effects of Kir6.2 mutations on channel gating. (A) Schematic of the different channel types expected when WT and mutant Kir6.2 are coexpressed (as in the heterozygous state). The box indicates channel types expected to have altered ATP sensitivity if the mutation affects channel gating (see text). (B) Single KATP channel currents recorded at –60 mV from inside-out patches from oocytes coinjected with mRNAs encoding SUR1 plus either WT or mutant Kir6.2 as indicated. Reproduced with permission from Proceedings of the National Academy of Sciences of the United States of America (83).
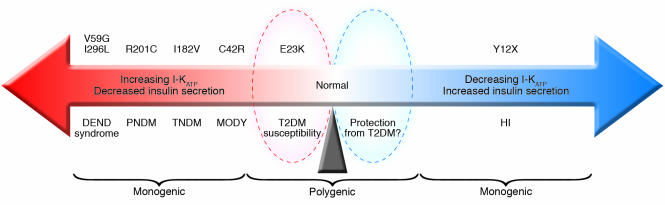
Schematic illustrating the relationship between KATP channel activity and insulin secretion. T2DM, type 2 diabetes mellitus.
References
-
- Ashcroft FM, Harrison DE, Ashcroft SJH. Glucose induces closure of single potassium channels in isolated rat pancreatic β-cells. Nature. 1984;312:446–448. - PubMed
-
- Koster JC, Marshall BA, Ensor N, Corbett JA, Nichols CG. Targeted overactivity of beta cell K(ATP) channels induces profound neonatal diabetes. Cell. 2000;100:645–654. - PubMed
-
- Seghers V, Nakazaki M, DeMayo F, Aguilar-Bryan L, Bryan J. Sur1 knockout mice. A model for K(ATP) channel-independent regulation of insulin secretion. J. Biol. Chem. 2000;275:9270–9277. - PubMed
