

Evolution and molecular phylogeny of Listeria monocytogenes isolated from human and animal listeriosis cases and foods
- PMID: 16077098
- PMCID: PMC1196091
- DOI: 10.1128/JB.187.16.5537-5551.2005
Evolution and molecular phylogeny of Listeria monocytogenes isolated from human and animal listeriosis cases and foods
Abstract
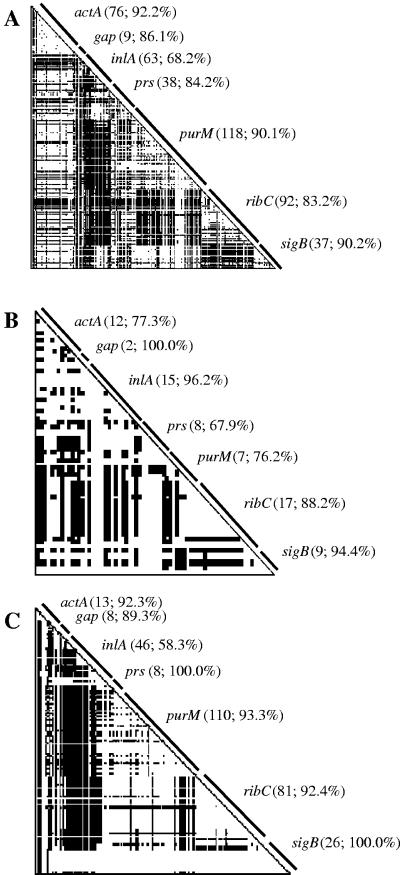
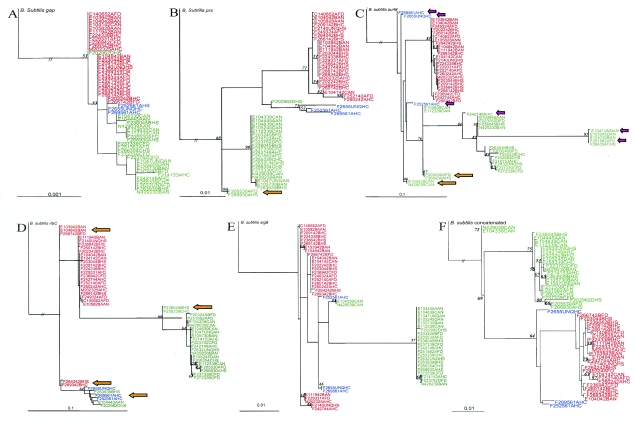
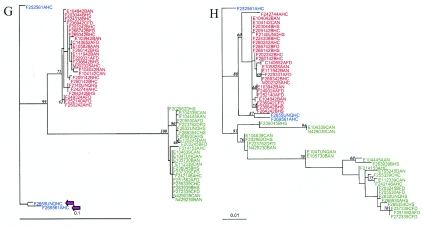
To probe the evolution and phylogeny of Listeria monocytogenes from defined host species and environments, L. monocytogenes isolates from human (n = 60) and animal (n = 30) listeriosis cases and food samples (n = 30) were randomly selected from a larger collection of isolates (n = 354) obtained in New York State between 1999 and 2001. Partial sequencing of four housekeeping genes (gap, prs, purM, and ribC), one stress response gene (sigB), and two virulence genes (actA and inlA) revealed between 11 (gap) and 33 (inlA) allelic types as well as 52 sequence types (unique combination of allelic types). actA, ribC, and purM demonstrated the highest levels of nucleotide diversity (pi > 0.05). actA and inlA as well as prs and the hypervariable housekeeping genes ribC and purM showed evidence of horizontal gene transfer and recombination. actA and inlA also showed evidence of positive selection at specific amino acid sites. Maximum likelihood phylogenies for all seven genes confirmed that L. monocytogenes contains two deeply separated evolutionary lineages. Lineage I was found to be highly clonal, while lineage II showed greater diversity and evidence of horizontal gene transfer. Allelic types were exclusive to lineages, except for a single gap allele, and nucleotide distance within lineages was much lower than that between lineages, suggesting that genetic exchange between lineages is rare. Our data show that (i) L. monocytogenes is a highly diverse species with at least two distinct phylogenetic lineages differing in their evolutionary history and population structure and (ii) horizontal gene transfer as well as positive selection contributed to the evolution of L. monocytogenes.
Figures
References
-
- Bart, A., C. Barnabe, M. Achtman, J. Dankert, A. van der Ende, and M. Tibayrenc. 2001. The population structure of Neisseria meningitidis serogroup A fits the predictions for clonality. Infect. Gen. Evol. 1:117-122. - PubMed
-
- Bromham, L., and D. Penny. 2003. The modern molecular clock. Nat. Rev. Genet. 3:216-224. - PubMed
-
- Czajka, J., N. Bsat, M. Piani, W. Russ, K. Sultana, M. Wiedmann, R. Whitaker, and C. A. Batt. 1993. Differentiation of Listeria monocytogenes and Listeria innocua by 16S rRNA genes and intraspecies discrimination of Listeria monocytogenes strains by random amplified polymorphic DNA polymorphisms. Appl. Environ. Microbiol. 59:304-308. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
