

The bacteriophage 434 repressor dimer preferentially undergoes autoproteolysis by an intramolecular mechanism
- PMID: 16077107
- PMCID: PMC1196080
- DOI: 10.1128/JB.187.16.5624-5630.2005
The bacteriophage 434 repressor dimer preferentially undergoes autoproteolysis by an intramolecular mechanism
Abstract
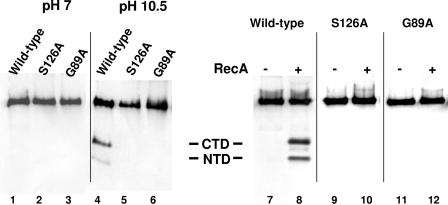
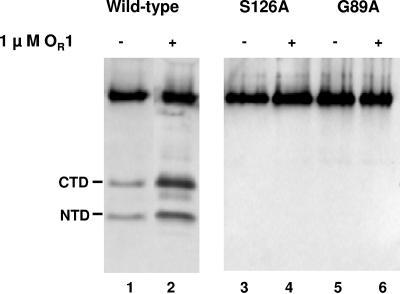
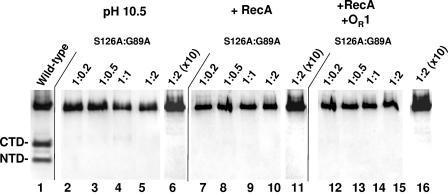
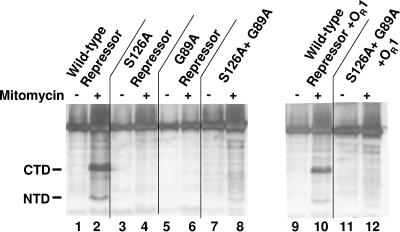
Inactivation of the lambdoid phage repressor protein is necessary to induce lytic growth of a lambdoid prophage. Activated RecA, the mediator of the host SOS response to DNA damage, causes inactivation of the repressor by stimulating the repressor's nascent autocleavage activity. The repressor of bacteriophage lambda and its homolog, LexA, preferentially undergo RecA-stimulated autocleavage as free monomers, which requires that each monomer mediates its own (intramolecular) cleavage. The cI repressor of bacteriophage 434 preferentially undergoes autocleavage as a dimer specifically bound to DNA, opening the possibility that one 434 repressor subunit may catalyze proteolysis of its partner subunit (intermolecular cleavage) in the DNA-bound dimer. Here, we first identified and mutagenized the residues at the cleavage and active sites of 434 repressor. We utilized the mutant repressors to show that the DNA-bound 434 repressor dimer overwhelmingly prefers to use an intramolecular mechanism of autocleavage. Our data suggest that the 434 repressor cannot be forced to use an intermolecular cleavage mechanism. Based on these data, we propose a model in which the cleavage-competent conformation of the repressor is stabilized by operator binding.
Figures

Similar articles
-
The preferred substrate for RecA-mediated cleavage of bacteriophage 434 repressor is the DNA-bound dimer.J Bacteriol. 2004 Jan;186(1):1-7. doi: 10.1128/JB.186.1.1-7.2004. J Bacteriol. 2004. PMID: 14679217 Free PMC article.
-
Carboxyl-terminal domain dimer interface mutant 434 repressors have altered dimerization and DNA binding specificities.J Mol Biol. 1998 Nov 13;283(5):931-46. doi: 10.1006/jmbi.1998.2136. J Mol Biol. 1998. PMID: 9799634
-
Purification and characterization of the repressor of the shiga toxin-encoding bacteriophage 933W: DNA binding, gene regulation, and autocleavage.J Bacteriol. 2004 Nov;186(22):7659-69. doi: 10.1128/JB.186.22.7659-7669.2004. J Bacteriol. 2004. PMID: 15516580 Free PMC article.
-
Regulation in repressor inactivation by RecA protein.Adv Biophys. 1990;26:33-49. doi: 10.1016/0065-227x(90)90006-f. Adv Biophys. 1990. PMID: 2082728 Review.
-
Cooperativity: action at a distance in a classic system.Curr Biol. 2000 Oct 5;10(19):R704-7. doi: 10.1016/s0960-9822(00)00710-7. Curr Biol. 2000. PMID: 11050405 Review.
Cited by
-
Noncanonical DNA-binding mode of repressor and its disassembly by antirepressor.Proc Natl Acad Sci U S A. 2016 May 3;113(18):E2480-8. doi: 10.1073/pnas.1602618113. Epub 2016 Apr 20. Proc Natl Acad Sci U S A. 2016. PMID: 27099293 Free PMC article.
-
Environmental stress perception activates structural remodeling of extant Streptococcus mutans biofilms.NPJ Biofilms Microbiomes. 2020 Mar 27;6(1):17. doi: 10.1038/s41522-020-0128-z. NPJ Biofilms Microbiomes. 2020. PMID: 32221309 Free PMC article.
-
Identification and characterization of the immunity repressor (ImmR) that controls the mobile genetic element ICEBs1 of Bacillus subtilis.Mol Microbiol. 2007 Jun;64(6):1515-28. doi: 10.1111/j.1365-2958.2007.05748.x. Epub 2007 May 18. Mol Microbiol. 2007. PMID: 17511812 Free PMC article.
References
-
- Bell, A. C., and G. B. Koudelka. 1993. Operator sequence context influences amino acid-base-pair interactions in 434 repressor-operator complexes. J. Mol. Biol. 234:542-553. - PubMed
-
- Bushman, F. D. 1993. The bacteriophage 434 right operator. Roles of OR1, OR2 and OR3. J. Mol. Biol. 230:28-40. - PubMed
-
- Ciubotaru, M., F. V. Bright, C. M. Ingersoll, and G. B. Koudelka. 1999. DNA-induced conformational changes in bacteriophage 434 repressor. J. Mol. Biol. 294:859-873. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
