

HepG2 cells develop signs of riboflavin deficiency within 4 days of culture in riboflavin-deficient medium
- PMID: 16081269
- PMCID: PMC1373781
- DOI: 10.1016/j.jnutbio.2005.03.006
HepG2 cells develop signs of riboflavin deficiency within 4 days of culture in riboflavin-deficient medium
Abstract
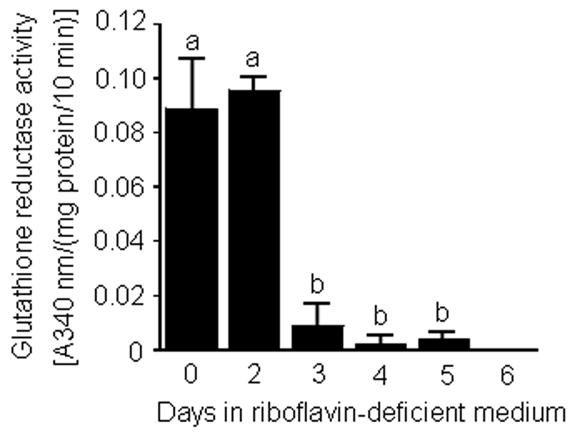
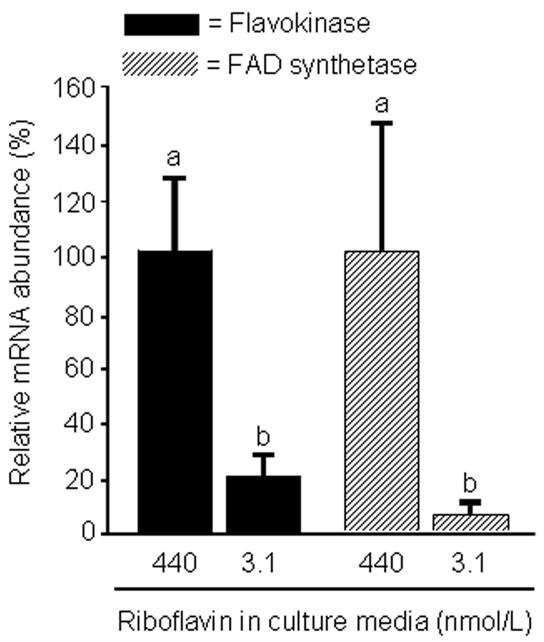
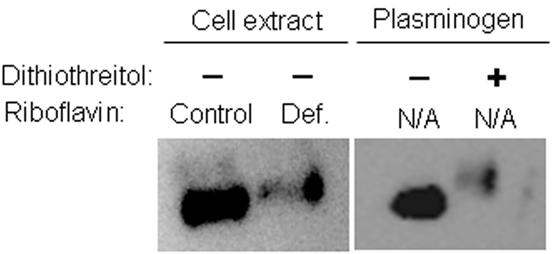
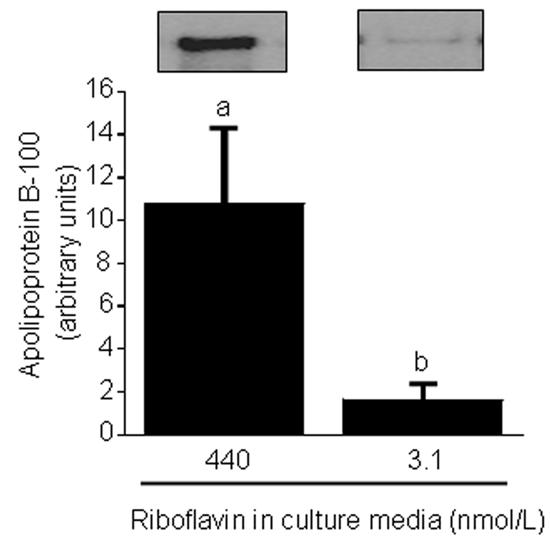
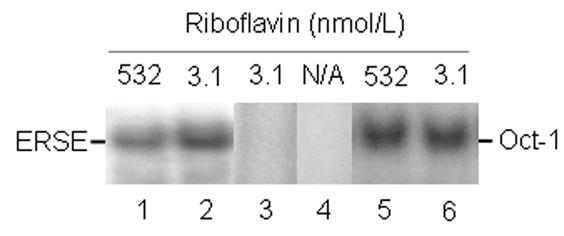
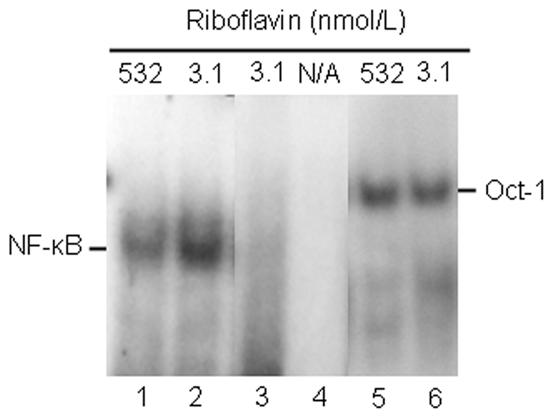
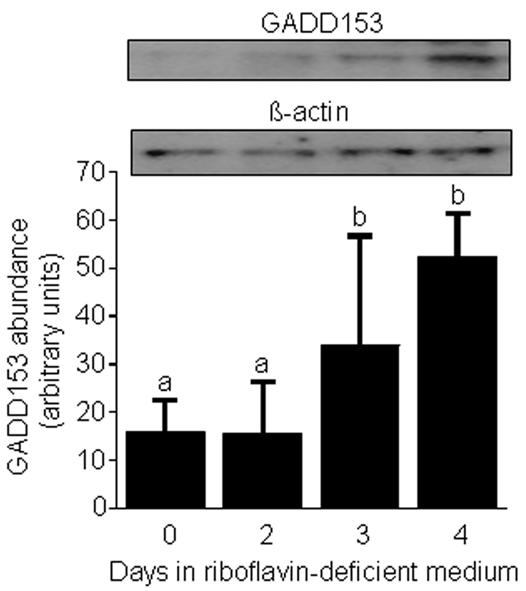
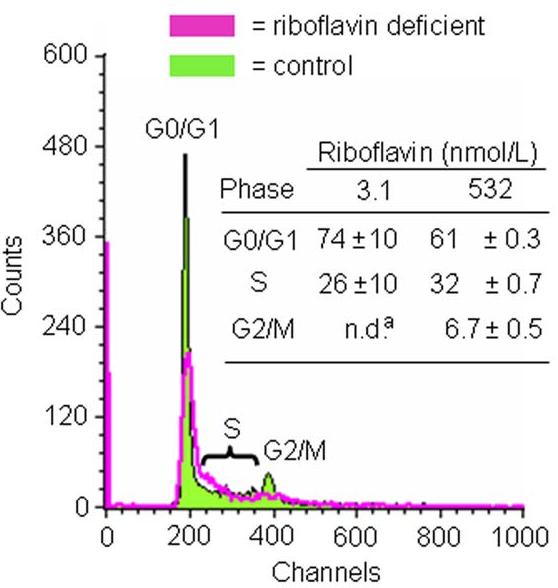
Flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD) are essential coenzymes in redox reactions. For example, FAD is a coenzyme for both glutathione reductase and enzymes that mediate the oxidative folding of secretory proteins. Here we investigated short-term effects of moderately riboflavin-deficient culture medium on flavin-related responses in HepG2 hepatocarcinoma cells. Cells were cultured in riboflavin-deficient (3.1 nmol/l) medium for up to 6 days; controls were cultured in riboflavin-sufficient (532 nmol/l) medium. The activity of glutathione reductase decreased by 98% within 4 days of riboflavin-deficient culture. Transport rates of riboflavin increased in response to riboflavin depletion, whereas expression of enzymes mediating flavocoenzyme synthesis (flavokinase and FAD synthetase) decreased in response to depletion. The oxidative folding and synthesis of plasminogen and apolipoprotein B-100 was impaired within 4 days of culture in riboflavin-deficient medium; this is consistent with impaired processing of secretory proteins in riboflavin-deficient cells. Riboflavin depletion was associated with increased DNA-binding activities of transcription factors with affinity for endoplasmic reticulum stress elements and nuclear factor kappaB (NF-kappaB) consensus elements, suggesting cell stress. Moreover, the abundance of the stress-induced protein GADD153 was greater in riboflavin-deficient cells compared with controls. Riboflavin deficiency was associated with decreased rates of cell proliferation caused by arrest in G1 phase of the cell cycle. These studies are consistent with the hypothesis that HepG2 cells have a great demand for riboflavin and that cell stress develops rapidly if riboflavin supply is marginally low.
Figures
References
-
- Rivlin RS. Riboflavin. In: Present Knowledge in Nutrition. In: Bowman BA, Russell RM, editors. ILSI Press; Washington, DC: 2001.
-
- Tu BP, Ho-Schleyer SC, Travers KL, Weissman JS. Biochemical basis of oxidative protein folding in the endoplasmic reticulum. Science. 2000;290:1571–1574. - PubMed
-
- Thorpe C, Hoober K, Raje S, Glynn N, Burnside J, Turi G, Coppock D. Sulfhydryl oxidases: emerging catalysts of protein disulfide bond formation in eukaryotes. Arch. Bioch. Biophys. 2002;405:1–12. - PubMed
-
- Camporeale G, Zempleni J. Oxidative folding of interleukin-2 is impaired in flavin-deficient Jurkat cells, causing intracellular accumulation of interleukin-2 and increased expression of stress response genes. J. Nutr. 2003;133:668–672. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
