

Intragenic tandem repeats generate functional variability
- PMID: 16086015
- PMCID: PMC1462868
- DOI: 10.1038/ng1618
Intragenic tandem repeats generate functional variability
Abstract
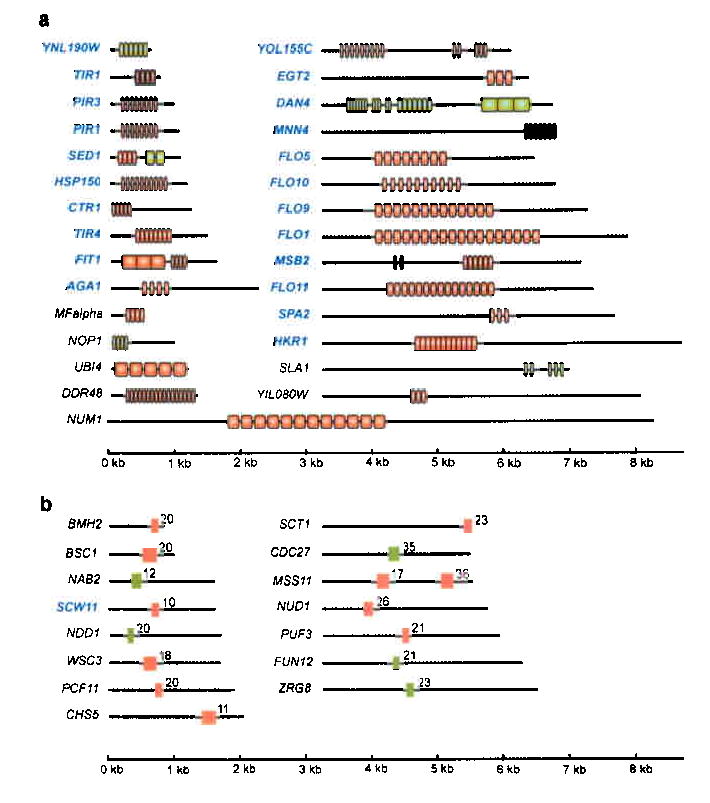
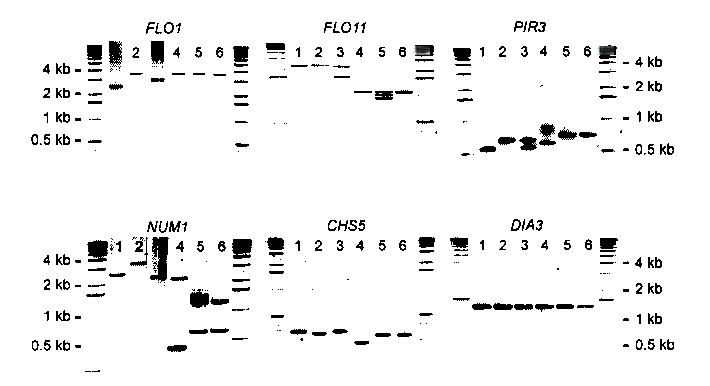
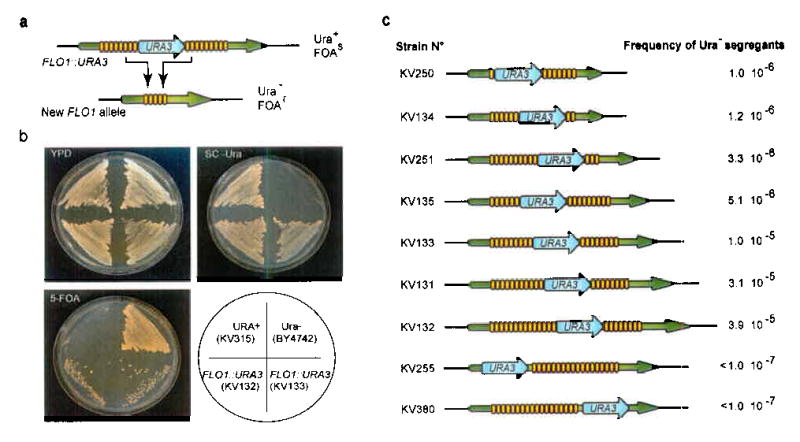
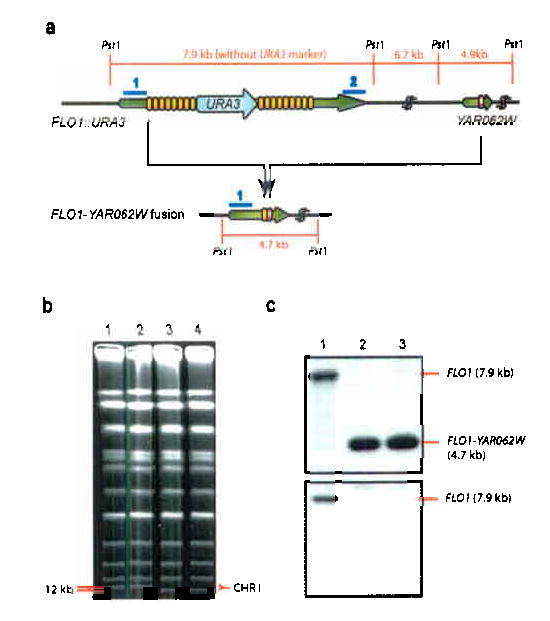
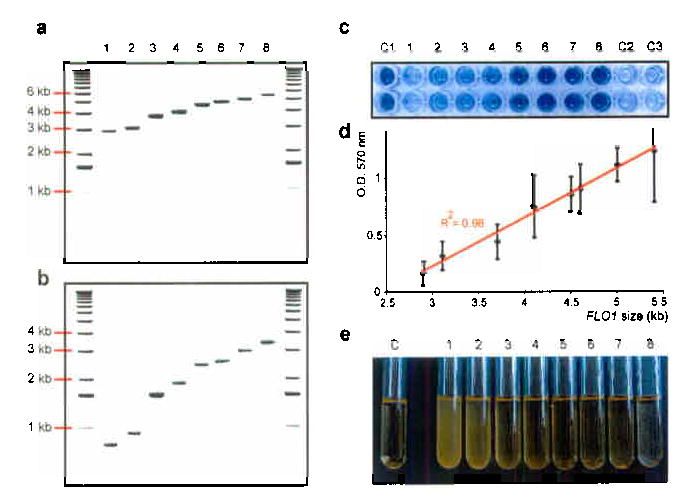
Tandemly repeated DNA sequences are highly dynamic components of genomes. Most repeats are in intergenic regions, but some are in coding sequences or pseudogenes. In humans, expansion of intragenic triplet repeats is associated with various diseases, including Huntington chorea and fragile X syndrome. The persistence of intragenic repeats in genomes suggests that there is a compensating benefit. Here we show that in the genome of Saccharomyces cerevisiae, most genes containing intragenic repeats encode cell-wall proteins. The repeats trigger frequent recombination events in the gene or between the gene and a pseudogene, causing expansion and contraction in the gene size. This size variation creates quantitative alterations in phenotypes (e.g., adhesion, flocculation or biofilm formation). We propose that variation in intragenic repeat number provides the functional diversity of cell surface antigens that, in fungi and other pathogens, allows rapid adaptation to the environment and elusion of the host immune system.
Figures
References
-
- Hartl DL. Molecular melodies in high and low C. Nature Rev Genet. 2000;1:145–149. - PubMed
-
- Verstrepen KJ, Reynolds TB, Fink GR. Origins of variation in the fungal cell surface. Nat Rev Microbiol. 2004;2:533–540. - PubMed
-
- Li YC, Korol AB, Fahima T, Nevo E. Microsatellites within genes: structure, function and evolution. Mol Biol Evol. 2004;21:991–1007. - PubMed
-
- Jin P, Alisch RS, Warren ST. RNA and microRNAs in fragile X mental retardation. Nat Cell Biol. 2004;6:1048–53. - PubMed
-
- Kellis M, Patterson N, Endrizzi M, Birren B, Lander ES. Sequencing and comparison of yeast species to identify genes and regulatory elements. Nature. 2003;423:241–54. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
