

Human T-cell lymphotropic virus type I-transformed T-cells have a partial defect in ceramide synthesis in response to N-(4-hydroxyphenyl)retinamide
- PMID: 16086670
- PMCID: PMC1317682
- DOI: 10.1042/BJ20050578
Human T-cell lymphotropic virus type I-transformed T-cells have a partial defect in ceramide synthesis in response to N-(4-hydroxyphenyl)retinamide
Erratum in
- Biochem J. 2012 Jun 15;444(3):619. Abou Tayyoun, Ahmad [corrected to Tayoun, Ahmad Abou]
Abstract
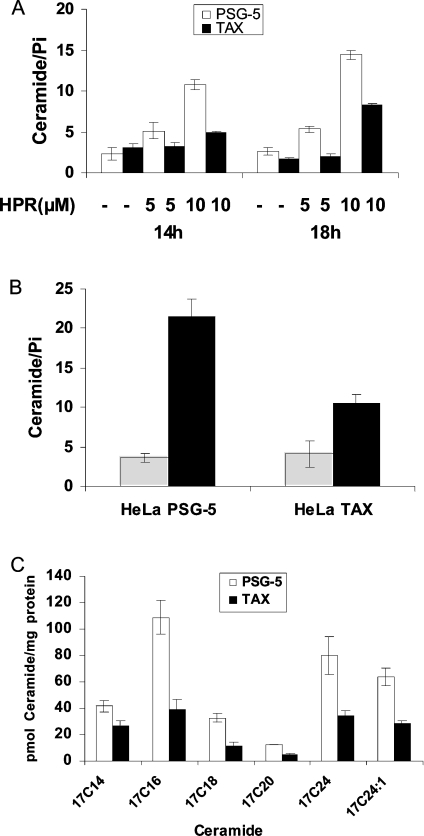
Treatment with the synthetic retinoid HPR [N-(4-hydroxyphenyl)-retinamide] causes growth arrest and apoptosis in HTLV-I (human T-cell lymphotropic virus type-I)-positive and HTLV-I-negative malignant T-cells. It was observed that HPR-mediated growth inhibition was associated with ceramide accumulation only in HTLV-I-negative cells. The aim of the present study was to investigate the mechanism by which HPR differentially regulates ceramide metabolism in HTLV-I-negative and HTLV-I-positive malignant T-cells. Clinically achievable concentrations of HPR caused early dose-dependent increases in ceramide levels only in HTLV-I-negative cells and preceded HPR-induced growth suppression. HPR induced de novo synthesis of ceramide in HTLV-I-negative, but not in HTLV-I-positive, cells. Blocking ceramide glucosylation in HTLV-I-positive cells, which leads to accumulation of endogenous ceramide, rendered these cells more sensitive to HPR. Exogenous cell-permeant ceramides that function partially by generating endogenous ceramide induced growth suppression in all tested malignant lymphocytes, were consistently found to be less effective in HTLV-I-positive cells confirming their defect in de novo ceramide synthesis. Owing to its multipotent activities, the HTLV-I-encoded Tax protein was suspected to inhibit ceramide synthesis. Tax-transfected Molt-4 and HELA cells were less sensitive to HPR and C6-ceramide mediated growth inhibition respectively and produced lower levels of endogenous ceramide. Together, these results indicate that HTLV-I-positive cells are defective in de novo synthesis of ceramide and that therapeutic modalities that bypass this defect are more likely to be successful.
Figures




Similar articles
-
N-(4-hydroxyphenyl)retinamide induces growth arrest and apoptosis in HTLV-I-transformed cells.Leukemia. 2004 Mar;18(3):607-15. doi: 10.1038/sj.leu.2403245. Leukemia. 2004. PMID: 14712289
-
Reactive oxygen species mediate N-(4-hydroxyphenyl)retinamide-induced cell death in malignant T cells and are inhibited by the HTLV-I oncoprotein Tax.Leukemia. 2007 Feb;21(2):261-9. doi: 10.1038/sj.leu.2404472. Epub 2006 Nov 23. Leukemia. 2007. PMID: 17122865
-
N-(4-hydroxyphenyl)retinamide increases ceramide and is cytotoxic to acute lymphoblastic leukemia cell lines, but not to non-malignant lymphocytes.Leukemia. 2002 May;16(5):902-10. doi: 10.1038/sj.leu.2402485. Leukemia. 2002. PMID: 11986953
-
N-(4-hydroxyphenyl)retinamide elevates ceramide in neuroblastoma cell lines by coordinate activation of serine palmitoyltransferase and ceramide synthase.Cancer Res. 2001 Jul 1;61(13):5102-5. Cancer Res. 2001. PMID: 11431347
-
Synergistic cytotoxicity in solid tumor cell lines between N-(4-hydroxyphenyl)retinamide and modulators of ceramide metabolism.J Natl Cancer Inst. 2000 Dec 6;92(23):1897-909. doi: 10.1093/jnci/92.23.1897. J Natl Cancer Inst. 2000. PMID: 11106681
Cited by
-
Dihydroceramide accumulation and reactive oxygen species are distinct and nonessential events in 4-HPR-mediated leukemia cell death.Biochem Cell Biol. 2012 Apr;90(2):209-23. doi: 10.1139/o2012-001. Epub 2012 Mar 19. Biochem Cell Biol. 2012. PMID: 22428532 Free PMC article.
-
Restoration of ceramide de novo synthesis by the synthetic retinoid ST1926 as it induces adult T-cell leukemia cell death.Biosci Rep. 2020 Oct 30;40(10):BSR20200050. doi: 10.1042/BSR20200050. Biosci Rep. 2020. PMID: 33048123 Free PMC article.
-
Ceramide Suppresses Influenza A Virus Replication In Vitro.J Virol. 2019 Mar 21;93(7):e00053-19. doi: 10.1128/JVI.00053-19. Print 2019 Apr 1. J Virol. 2019. PMID: 30700605 Free PMC article.
References
-
- Delia D., Aiello A., Lombardi L., Pelicci P. G., Grignani F., Grignani F., Formelli F., Menard S., Costa A., Veronesi U. N-(4-hydroxyphenyl)retinamide induces apoptosis of malignant hemopoietic cell lines including those unresponsive to retinoic acid. Cancer Res. 1993;53:6036–6041. - PubMed
-
- Wu J. M., DiPietrantonio A. M., Hsieh T. C. Mechanism of fenretinide (4-HPR)-induced cell death. Apoptosis. 2001;6:377–388. - PubMed
-
- Kalemkerian G. P., Slusher R., Ramalingam S., Gadgeel S., Mabry M. Growth inhibition and induction of apoptosis by fenretinide in small-cell lung cancer cell lines. J. Natl. Cancer Inst. 1995;87:1674–1680. - PubMed
-
- Maurer B. J., Metelitsa L. S., Seeger R. C., Cabot M. C., Reynolds C. P. Increase of ceramide and induction of mixed apoptosis/necrosis by N-(4-hydroxyphenyl)-retinamide in neuroblastoma cell lines. J. Natl. Cancer Inst. 1999;91:1138–1146. - PubMed
-
- Oridate N., Suzuki S., Higuchi M., Mitchell M. F., Hong W. K., Lotan R. Involvement of reactive oxygen species in N-(4-hydroxyphenyl)retinamide-induced apoptosis in cervical carcinoma cells. J. Natl. Cancer Inst. 1997;89:1191–1198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources