

IL-4 receptor expression on CD8+ T cells is required for the development of protective memory responses against liver stages of malaria parasites
- PMID: 16087712
- PMCID: PMC2212849
- DOI: 10.1084/jem.20042463
IL-4 receptor expression on CD8+ T cells is required for the development of protective memory responses against liver stages of malaria parasites
Abstract
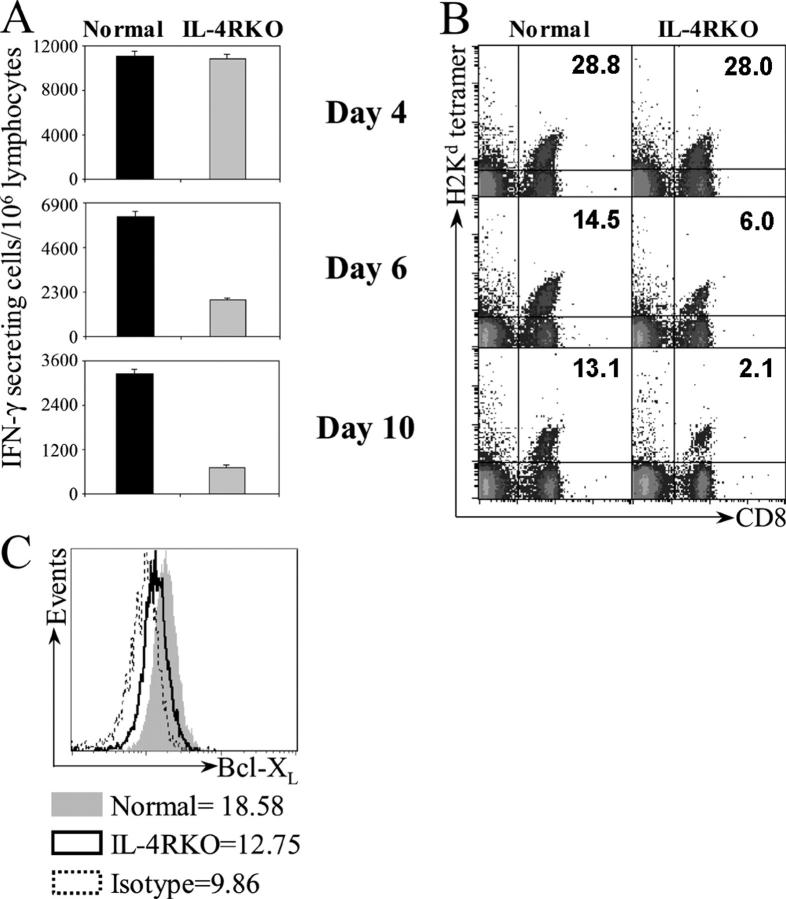
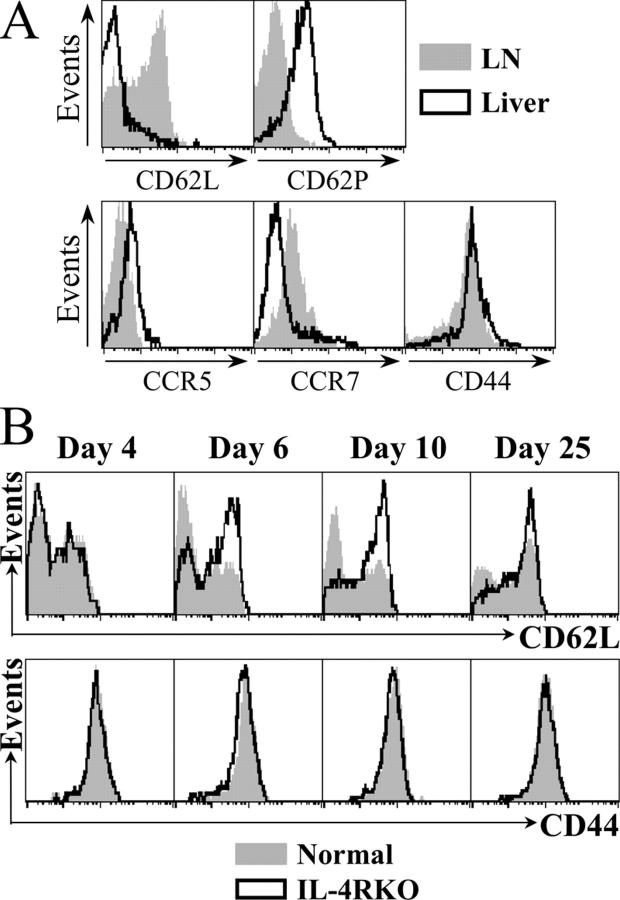
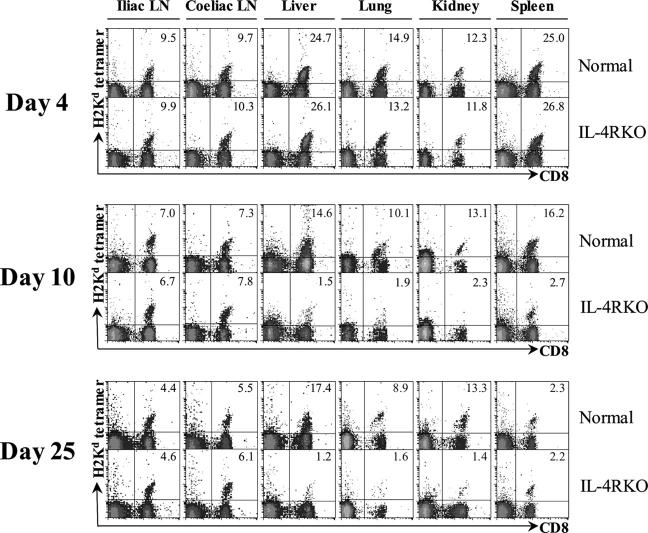
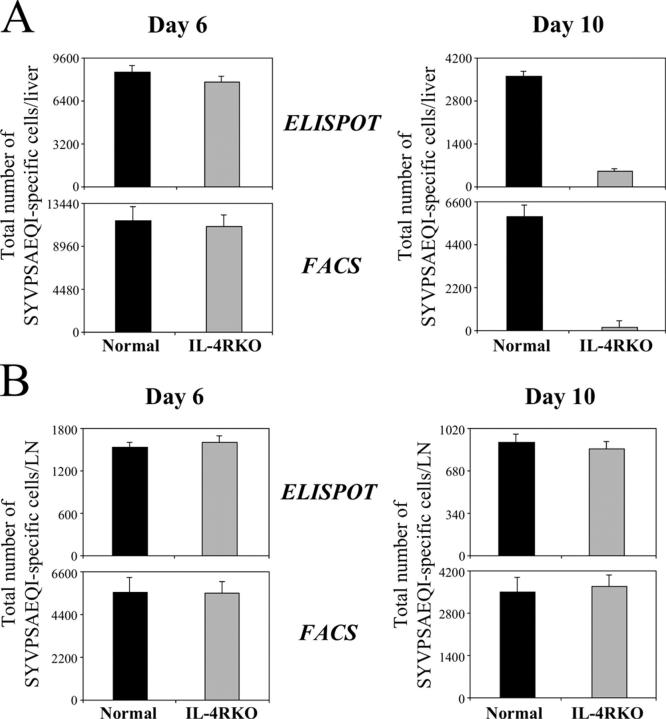
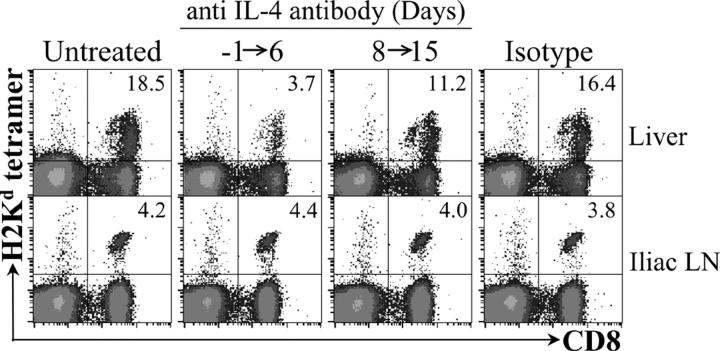
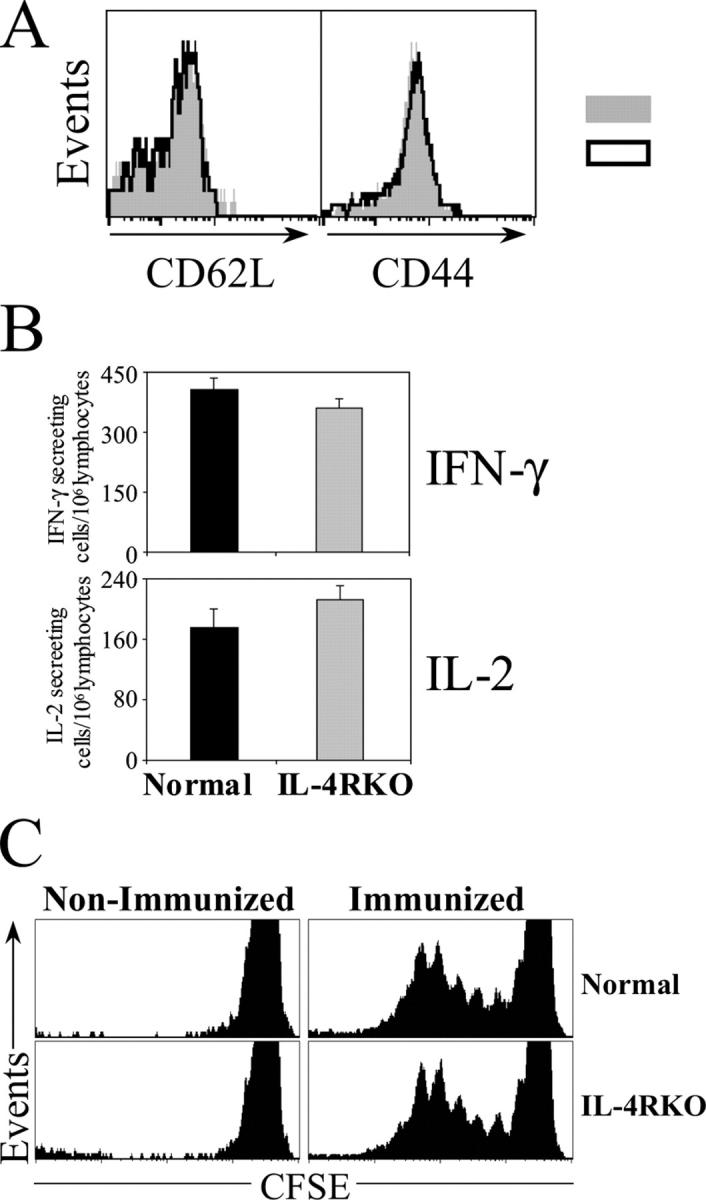
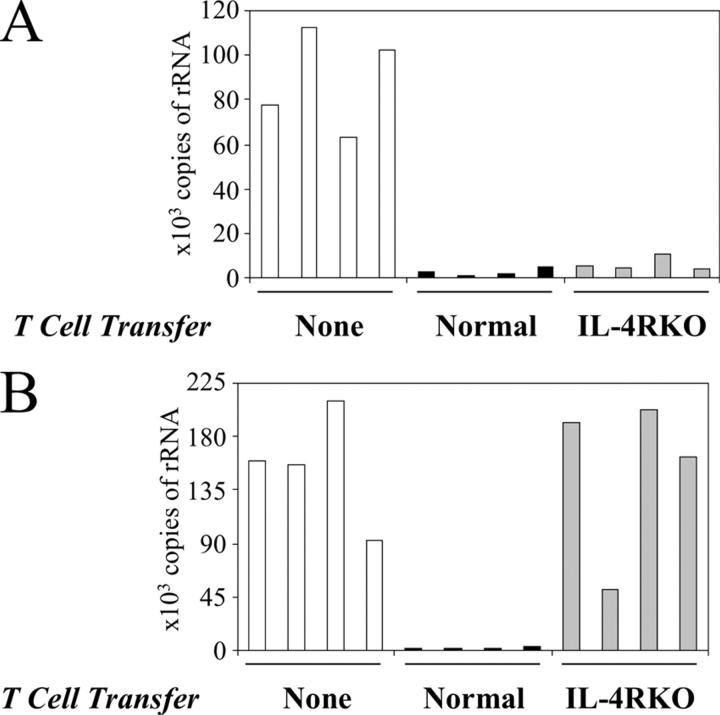
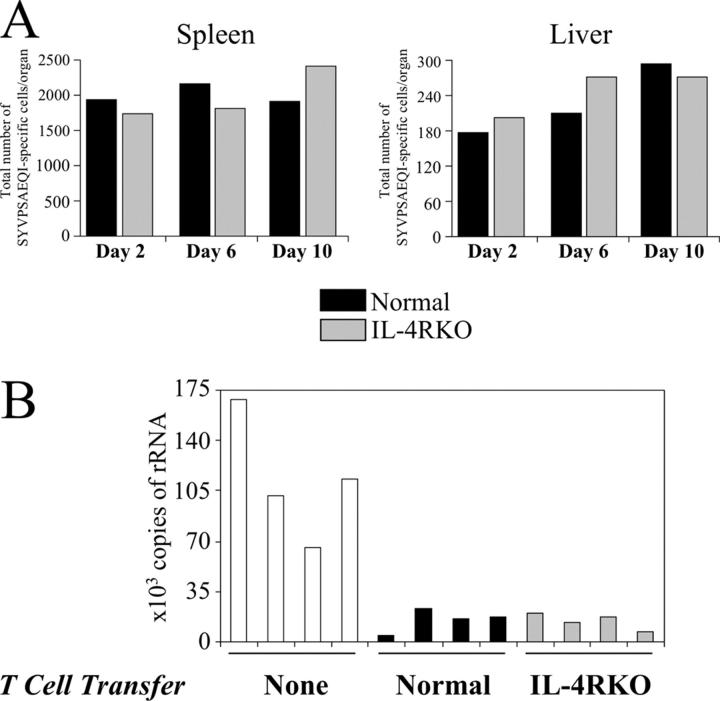
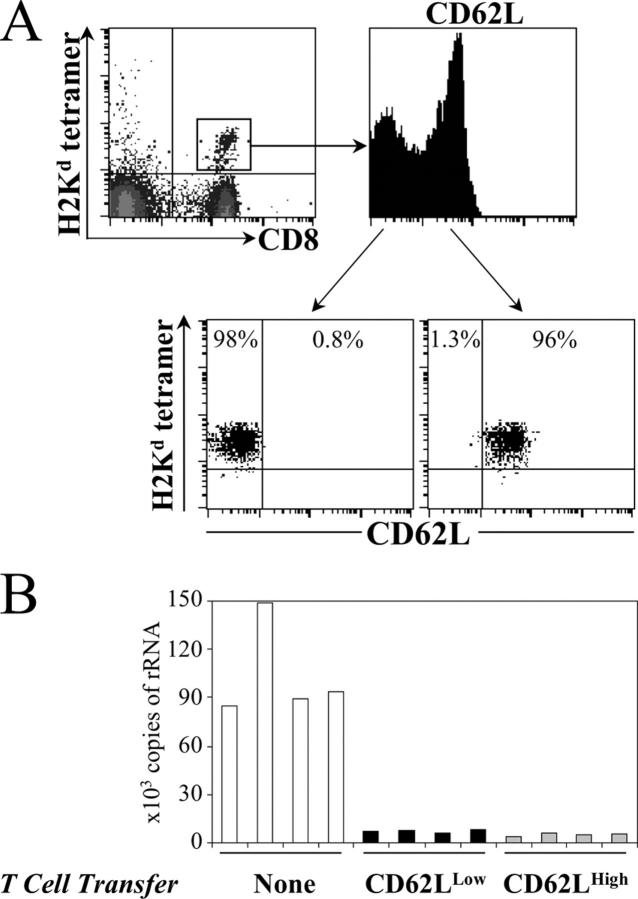
IL-4 receptor (IL-4R)-deficient CD8+ T cells specific for the circumsporozoite protein of Plasmodium yoelii develop a severely impaired memory response after priming with parasites. Memory CD8+ T cells lacking the IL-4R are unable to establish a stable population residing in nonlymphoid organs, although they develop normally in lymphoid organs. Because memory cells from nonlymphoid organs disappear shortly after immunization, the protective antiparasitic activity of this T cell response also is lost. These results demonstrate that IL-4/IL-4R interactions on CD8+ T cells play a critical role in modulating the development and tissue distribution of memory cells induced by parasite immunization. They also indicate that memory cells residing in nonlymphoid tissues are critical for protective immunity against malaria parasites.
Figures
References
-
- Wong, P., and E.G.P. Am. 2003. CD8 T cell responses to infectious pathogens. Annu. Rev. Immunol. 21:29–70. - PubMed
-
- Mueller, S.N., C.M. Jones, C.M. Smith, W.R. Heath, and F.R. Carbone. 2002. Rapid cytotoxic T lymphocyte activation occurs in the draining lymph nodes after cutaneous herpes simplex virus infection as a result of early antigen presentation and not the presence of virus. J. Exp. Med. 195:651–656. - PMC - PubMed
-
- Kaech, S.M., S. Hemby, E. Kersh, and R. Ahmed. 2002. Molecular and functional profiling of memory CD8 T cell differentiation. Cell. 111:837–851. - PubMed
-
- Lanzavecchia, A., and F. Sallusto. 2002. Progressive differentiation and selection of the fittest in the immune response. Nat. Rev. Immunol. 2:982–987. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
