

Identification and analysis of ribonuclease P and MRP RNA in a broad range of eukaryotes
- PMID: 16087735
- PMCID: PMC1183490
- DOI: 10.1093/nar/gki756
Identification and analysis of ribonuclease P and MRP RNA in a broad range of eukaryotes
Abstract
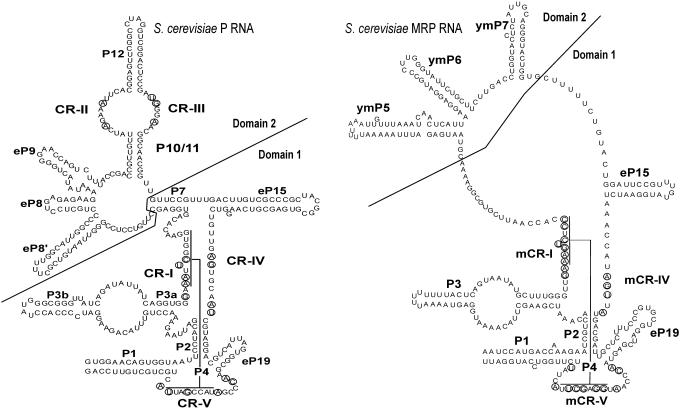
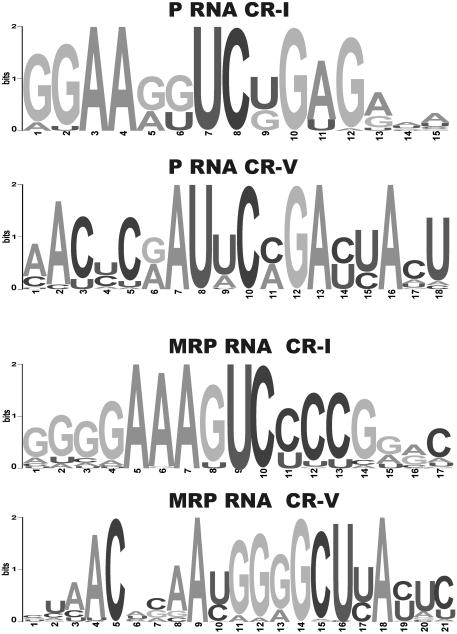
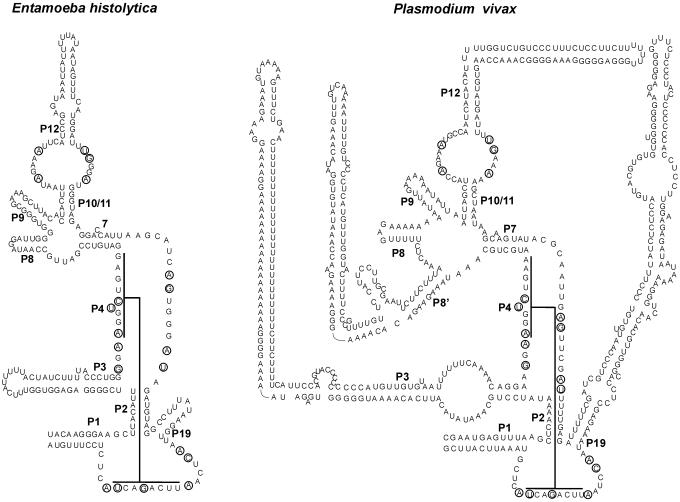
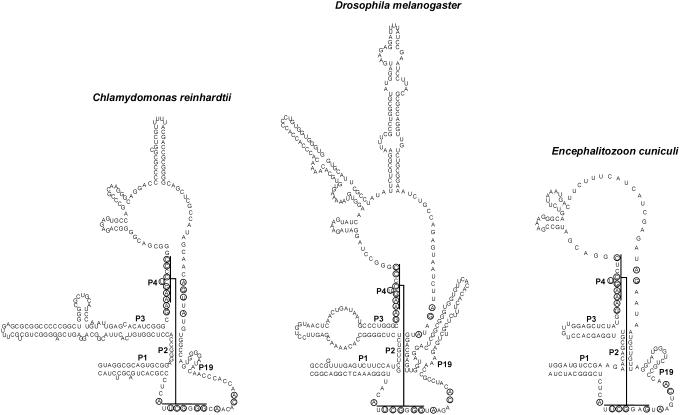
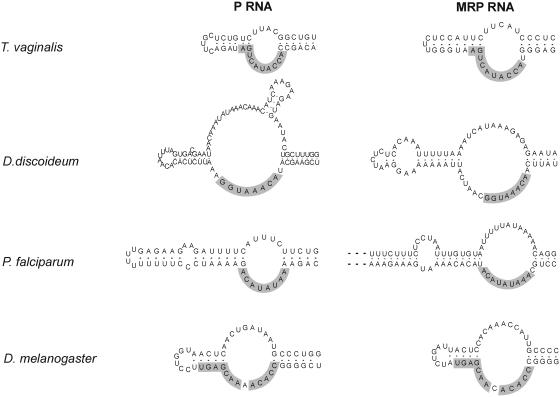
RNases P and MRP are ribonucleoprotein complexes involved in tRNA and rRNA processing, respectively. The RNA subunits of these two enzymes are structurally related to each other and play an essential role in the enzymatic reaction. Both of the RNAs have a highly conserved helical region, P4, which is important in the catalytic reaction. We have used a bioinformatics approach based on conserved elements to computationally analyze available genomic sequences of eukaryotic organisms and have identified a large number of novel nuclear RNase P and MRP RNA genes. For MRP RNA for instance, this investigation increases the number of known sequences by a factor of three. We present secondary structure models of many of the predicted RNAs. Although all sequences are able to fold into the consensus secondary structure of P and MRP RNAs, a striking variation in size is observed, ranging from a Nosema locustae MRP RNA of 160 nt to much larger RNAs, e.g. a Plasmodium knowlesi P RNA of 696 nt. The P and MRP RNA genes appear in tandem in some protists, further emphasizing the close evolutionary relationship of these RNAs.
Figures

References
-
- Lygerou Z., Mitchell P., Petfalski E., Seraphin B., Tollervey D. The POP1 gene encodes a protein component common to the RNase MRP and RNase P ribonucleoproteins. Genes Dev. 1994;8:1423–1433. - PubMed
-
- Lygerou Z., Allmang C., Tollervey D., Seraphin B. Accurate processing of a eukaryotic precursor ribosomal RNA by ribonuclease MRP in vitro. Science. 1996;272:268–270. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
