

Hydrogen exchange measurement of the free energy of structural and allosteric change in hemoglobin
- PMID: 1609279
- PMCID: PMC3562468
- DOI: 10.1126/science.256.5064.1684
Hydrogen exchange measurement of the free energy of structural and allosteric change in hemoglobin
Abstract
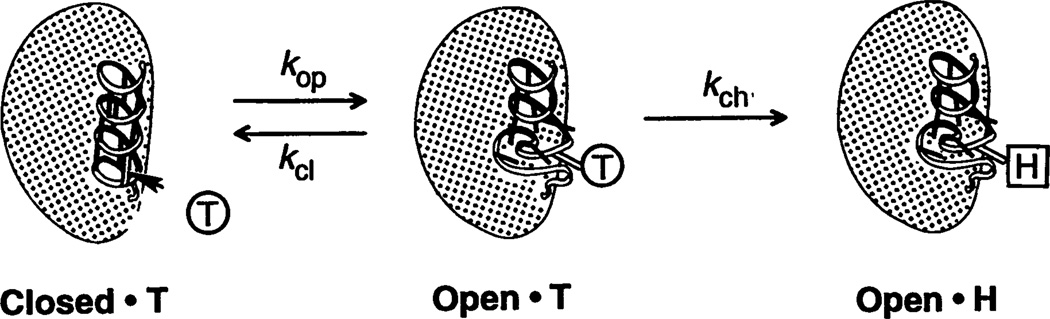
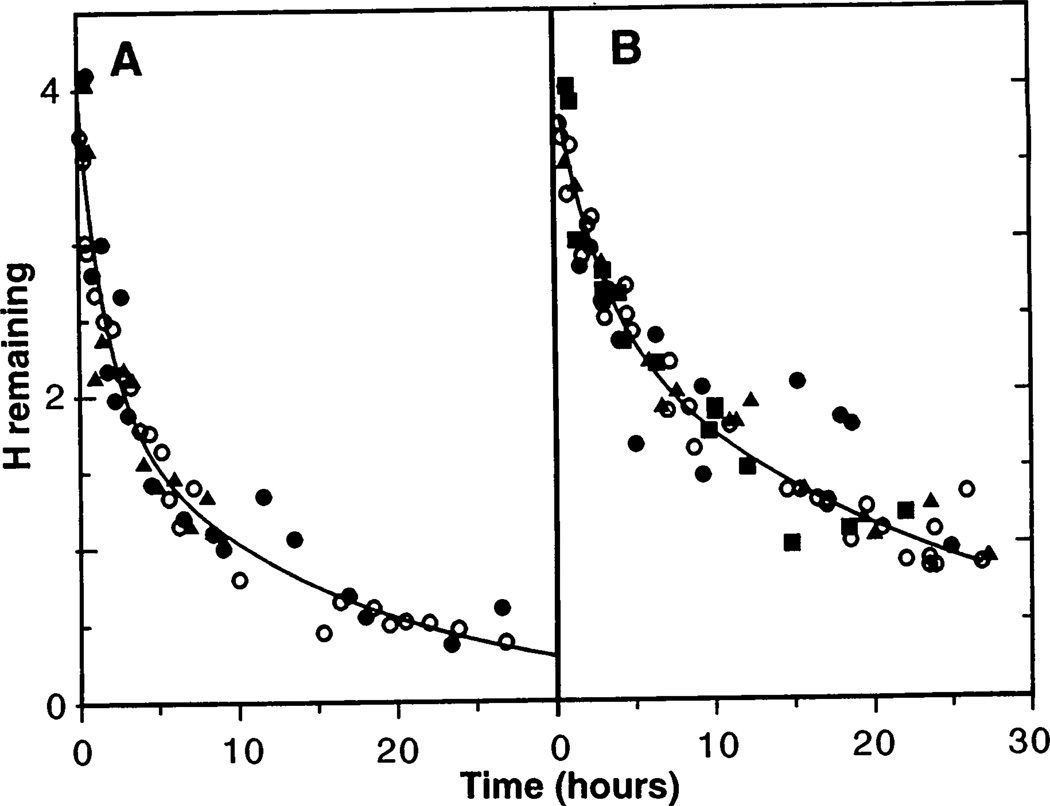
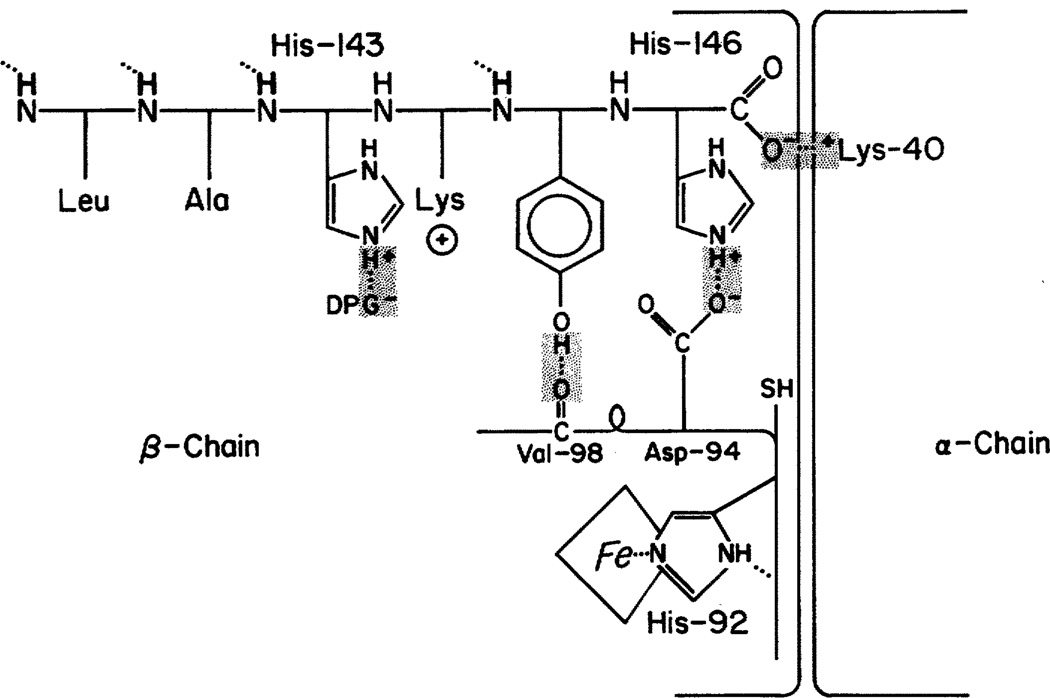
The inability to localize and measure the free energy of protein structure and structure change severely limits protein structure-function investigations. The local unfolding model for protein hydrogen exchange quantitatively related the free energy of local structural stability with the hydrogen exchange rate of concerted sets of structurally related protons. In tests with a number of modified hemoglobin forms, the loss in structural free energy obtained locally from hydrogen exchange results matches the loss in allosteric free energy measured globally by oxygen-binding and subunit dissociation experiments.
Figures
References
-
- Englander SW. Ann. N.Y Acad. Sci. 1975;244:10. - PubMed
-
- Englander JJ, Rogero JR, Englander SW. J. Mol. Biol. 1983;169:325. - PubMed
-
- Englander SW, Englander JJ. In: Structure and Dynamics of Nucleic Acids and Proteins. Clementi E, Sarma RH, editors. Guilderland, NY: Adenine; 1983. pp. 421–434.
- Rogero JR, Englander SW, Englander JJ. Methods Enzymol. 1986;131:L508. - PubMed
- Ray J, Englander SW. Biochemistry. 1986;25:3000. - PubMed
- Louie G, Tran T, Englander JJ, Englander SW. J. Mot. Biol. 1988;201:755. - PubMed
- Louie G, Englander JJ, Englander SW. ibid. :765. - PubMed
- McKinnie RE, Englander JJ, Englander SW. Chem. Phys. 1991;158:283.
-
-
Functional labeling is now generically termed hydrogen exchange labeling in parallel with analogous methods being used to study kinetic refolding intermediates, equilibrium unfolding intermediates, and protein interaction sites. For a review, see Englander SW, Mayne L. Annu. Rev. Biophys. Biomol. Struct. 1992;21:243.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
