

Deciphering the molecular bases of Mycobacterium tuberculosis binding to the lectin DC-SIGN reveals an underestimated complexity
- PMID: 16092920
- PMCID: PMC1316302
- DOI: 10.1042/BJ20050709
Deciphering the molecular bases of Mycobacterium tuberculosis binding to the lectin DC-SIGN reveals an underestimated complexity
Abstract
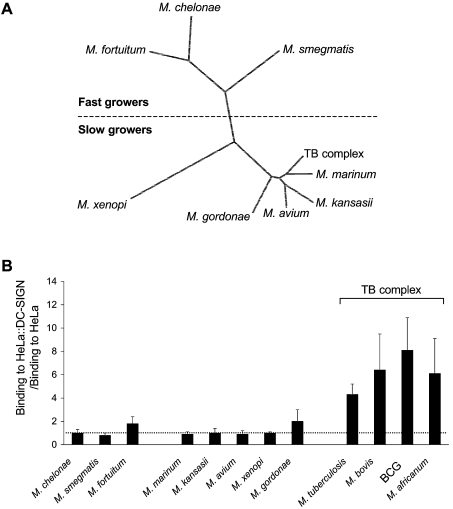
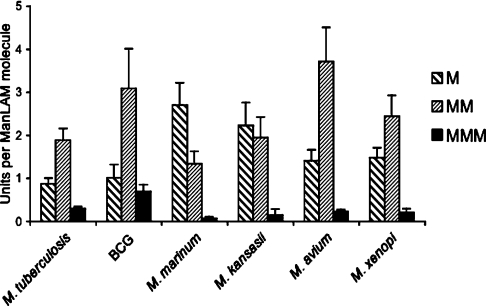
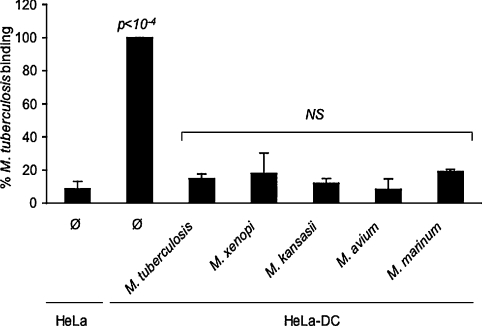
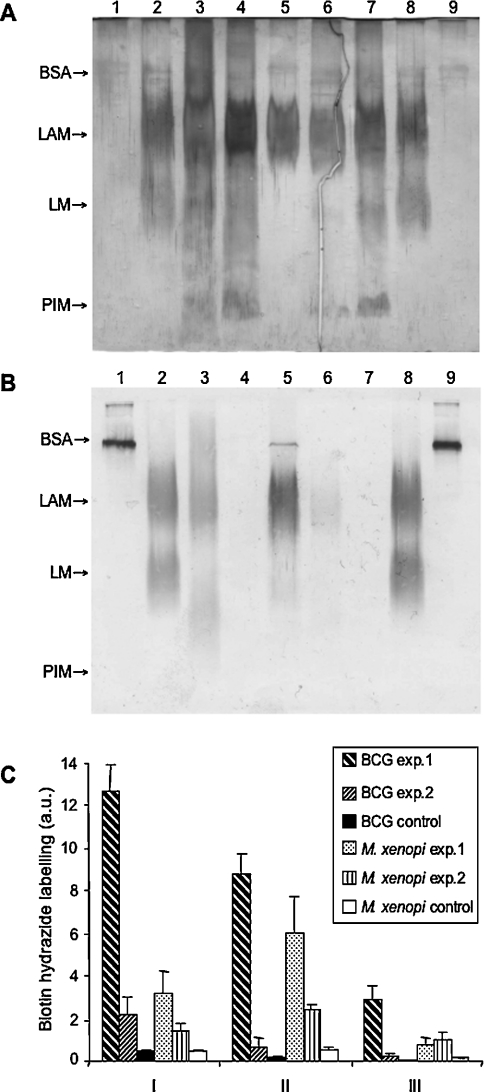
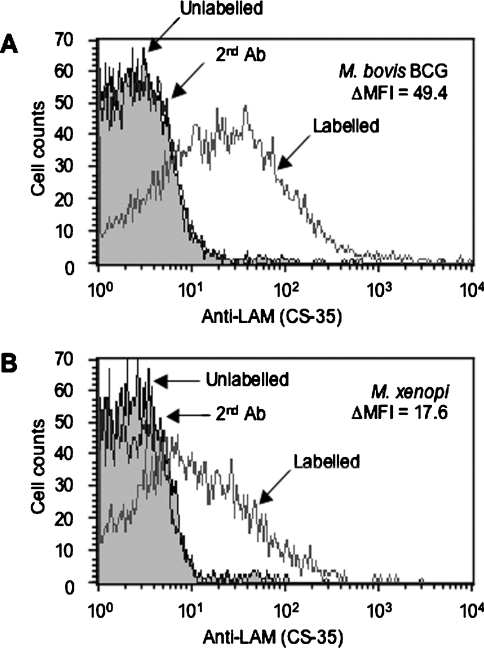
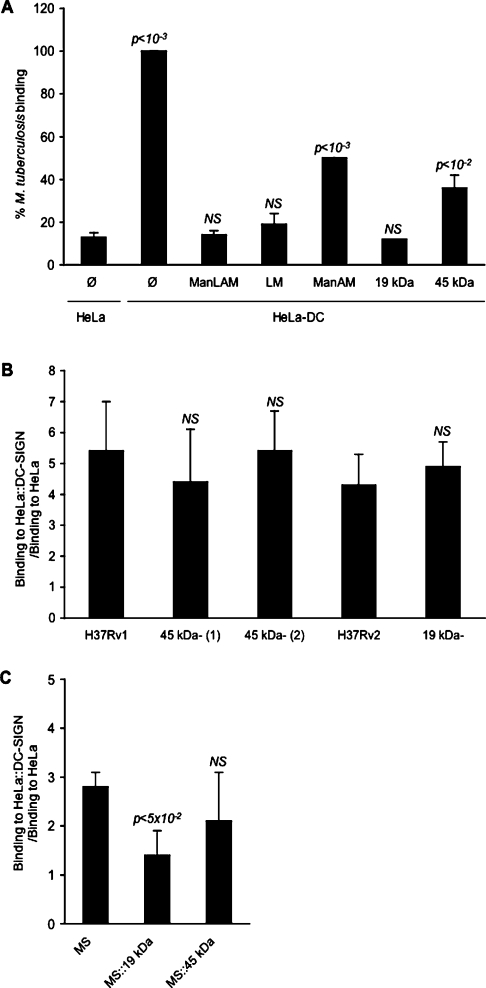
Interactions between dendritic cells and Mycobacterium tuberculosis, the aetiological agent of tuberculosis in humans, are thought to be central to anti-mycobacterial immunity. We have previously shown that M. tuberculosis binds to human monocyte-derived dendritic cells mostly through the C-type lectin DC-SIGN (dendritic-cell-specific intercellular molecule-3-grabbing non-integrin)/CD209, and we have suggested that DC-SIGN may discriminate between mycobacterial species through recognition of the mannose-capping residues on the lipoglycan lipoarabinomannan of the bacterial envelope. Here, using a variety of fast- and slow-growing Mycobacterium species, we provide further evidence that mycobacteria recognition by DC-SIGN may be restricted to species of the M. tuberculosis complex. Fine analyses of the lipoarabinomannan molecules purified from these species show that the structure and amount of these molecules alone cannot account for such a preferential recognition. We propose that M. tuberculosis recognition by DC-SIGN relies on both a potential difference of accessibility of lipoarabinomannan in its envelope and, more probably, on the binding of additional ligands, possibly including lipomannan, mannose-capped arabinomannan, as well as the mannosylated 19 kDa and 45 kDa [Apa (alanine/proline-rich antigen)] glycoproteins. Altogether, our results reveal that the molecular basis of M. tuberculosis binding to DC-SIGN is more complicated than previously thought and provides further insight into the mechanisms of M. tuberculosis recognition by the immune system.
Figures
Similar articles
-
Identification of four novel DC-SIGN ligands on Mycobacterium bovis BCG.Protein Cell. 2010 Sep;1(9):859-70. doi: 10.1007/s13238-010-0101-3. Epub 2010 Oct 7. Protein Cell. 2010. PMID: 21203928 Free PMC article.
-
The cell surface receptor DC-SIGN discriminates between Mycobacterium species through selective recognition of the mannose caps on lipoarabinomannan.J Biol Chem. 2003 Feb 21;278(8):5513-6. doi: 10.1074/jbc.C200586200. Epub 2002 Dec 20. J Biol Chem. 2003. PMID: 12496255
-
DC-SIGN is the major Mycobacterium tuberculosis receptor on human dendritic cells.J Exp Med. 2003 Jan 6;197(1):121-7. doi: 10.1084/jem.20021468. J Exp Med. 2003. PMID: 12515819 Free PMC article.
-
DC-SIGN and mannosylated surface structures of Mycobacterium tuberculosis: a deceptive liaison.Eur J Cell Biol. 2010 Jan;89(1):95-101. doi: 10.1016/j.ejcb.2009.10.004. Epub 2009 Nov 4. Eur J Cell Biol. 2010. PMID: 19892432 Review.
-
A fatal attraction: Mycobacterium tuberculosis and HIV-1 target DC-SIGN to escape immune surveillance.Trends Mol Med. 2003 Apr;9(4):153-9. doi: 10.1016/s1471-4914(03)00027-3. Trends Mol Med. 2003. PMID: 12727141 Review.
Cited by
-
DC-SIGN induction in alveolar macrophages defines privileged target host cells for mycobacteria in patients with tuberculosis.PLoS Med. 2005 Dec;2(12):e381. doi: 10.1371/journal.pmed.0020381. Epub 2005 Nov 15. PLoS Med. 2005. PMID: 16279841 Free PMC article.
-
Identification of four novel DC-SIGN ligands on Mycobacterium bovis BCG.Protein Cell. 2010 Sep;1(9):859-70. doi: 10.1007/s13238-010-0101-3. Epub 2010 Oct 7. Protein Cell. 2010. PMID: 21203928 Free PMC article.
-
Nanoscale clustering of mycobacterial ligands and DC-SIGN host receptors are key determinants for pathogen recognition.Sci Adv. 2023 May 19;9(20):eadf9498. doi: 10.1126/sciadv.adf9498. Epub 2023 May 19. Sci Adv. 2023. PMID: 37205764 Free PMC article.
-
C-type lectin DC-SIGN: an adhesion, signalling and antigen-uptake molecule that guides dendritic cells in immunity.Cell Signal. 2010 Oct;22(10):1397-405. doi: 10.1016/j.cellsig.2010.03.018. Epub 2010 Apr 2. Cell Signal. 2010. PMID: 20363321 Free PMC article. Review.
-
Scrutiny of Mycobacterium tuberculosis 19 kDa antigen proteoforms provides new insights in the lipoglycoprotein biogenesis paradigm.Sci Rep. 2017 Mar 8;7:43682. doi: 10.1038/srep43682. Sci Rep. 2017. PMID: 28272507 Free PMC article.
References
-
- Kaufmann S. H. How can immunology contribute to the control of tuberculosis? Nat. Rev. Immunol. 2001;1:20–30. - PubMed
-
- Flynn J. L., Chan J. Immunology of tuberculosis. Annu. Rev. Immunol. 2001;9:93–129. - PubMed
-
- Geijtenbeek T. B., Torensma R., van Vliet S. J., van Duijnhoven G. C., Adema G. J., van Kooyk Y., Figdor C. G. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell. 2000;100:575–585. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases