

Brain and muscle Arnt-like protein-1 (BMAL1), a component of the molecular clock, regulates adipogenesis
- PMID: 16093318
- PMCID: PMC1189312
- DOI: 10.1073/pnas.0502383102
Brain and muscle Arnt-like protein-1 (BMAL1), a component of the molecular clock, regulates adipogenesis
Abstract
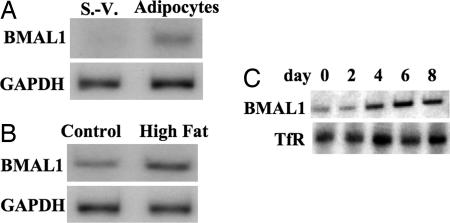
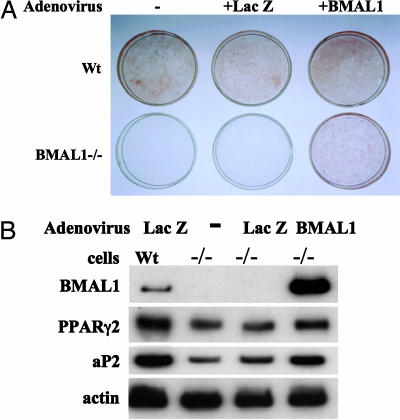
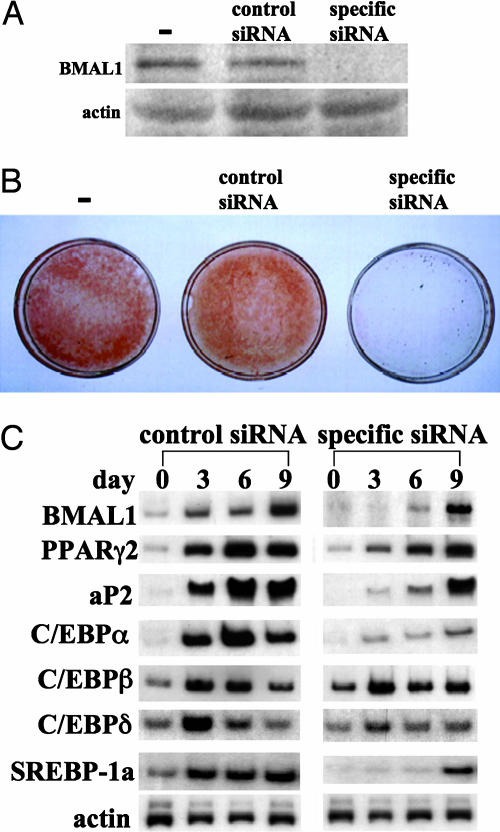
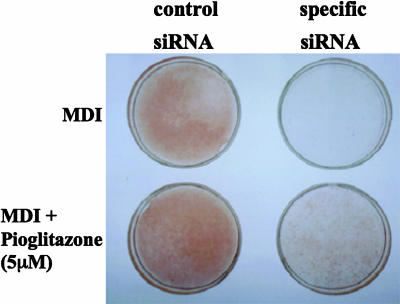
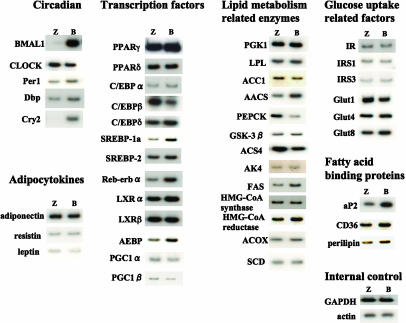
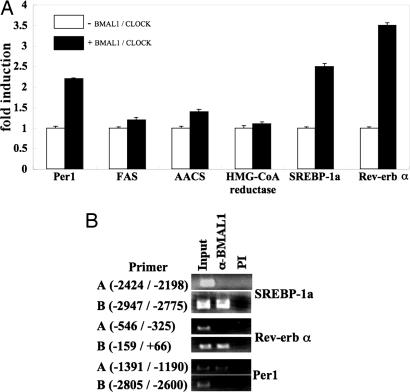
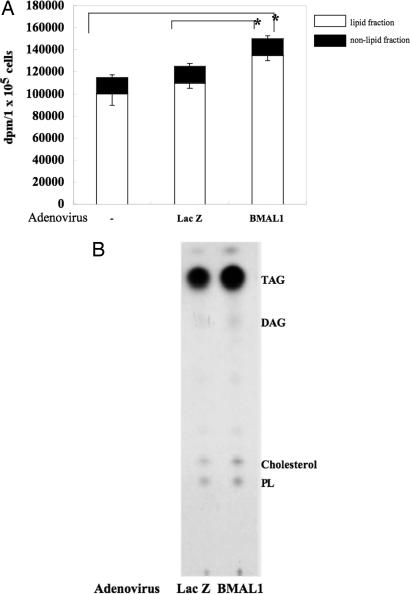
Brain and muscle Arnt-like protein-1 (BMAL1; also known as MOP3 or Arnt3) is a transcription factor known to regulate circadian rhythm. Here, we established its involvement in the control of adipogenesis and lipid metabolism activity in mature adipocytes. During adipose differentiation in 3T3-L1 cells, the level of BMAL1 mRNA began to increase 4 days after induction and was highly expressed in differentiated cells. In white adipose tissues isolated from C57BL/6J mice, BMAL1 was predominantly expressed in a fraction containing adipocytes, as compared with the stromal-vascular fraction. BMAL1 knockout mice embryonic fibroblast cells failed to be differentiated into adipocytes. Importantly, adding BMAL1 back by adenovirus gene transfer restored the ability of BMAL1 knockout mice embryonic fibroblast cells to differentiate. Knock-down of BMAL1 expression in 3T3-L1 cells by an RNA interference technique allowed the cells to accumulate only minimum amounts of lipid droplets in the cells. Adenovirus-mediated expression of BMAL1 in 3T3-L1 adipocytes resulted in induction of several factors involved in lipogenesis. The promoter activity of these genes was stimulated in a BMAL1-dependent manner. Interestingly, expression of these factors showed clear circadian rhythm in mice adipose tissue. Furthermore, overexpression of BMAL1 in adipocytes increased lipid synthesis activity. These results indicate that BMAL1, a master regulator of circadian rhythm, also plays important roles in the regulation of adipose differentiation and lipogenesis in mature adipocytes.
Figures
References
-
- Spiegelman, B. M. & Flier, J. S. (2001) Cell 104, 531–543. - PubMed
-
- Ahima, R. S. & Flier, J. S. (2000) Trends Endocrinol. Metab. 11, 327–332. - PubMed
-
- Seaman, G. V., Engel, R., Swank, R. I. & Hissen, W. (1965) Nature 207, 833–835. - PubMed
-
- Malherbe, C., De Gasparo, M., De Hertogh, R. & Hoet, J. J. (1969) Diabetologia 5, 397–404. - PubMed
-
- Gagliardino, J. J. & Hernandez, R. E. (1971) Endocrinology 88, 1532–1534. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
