

Prefrontal control of the amygdala
- PMID: 16093394
- PMCID: PMC6725290
- DOI: 10.1523/JNEUROSCI.2314-05.2005
Prefrontal control of the amygdala
Abstract
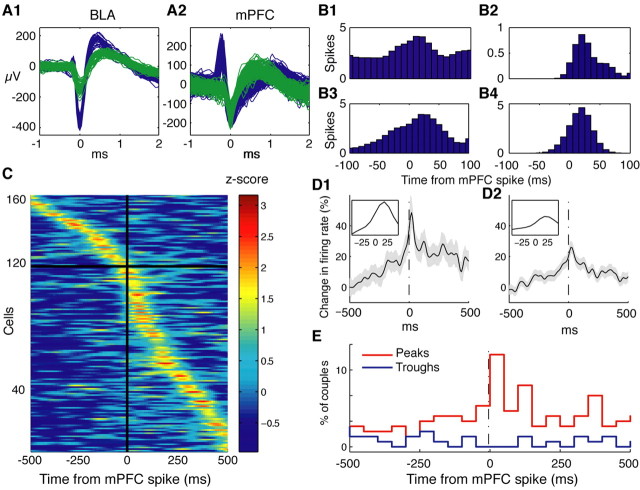
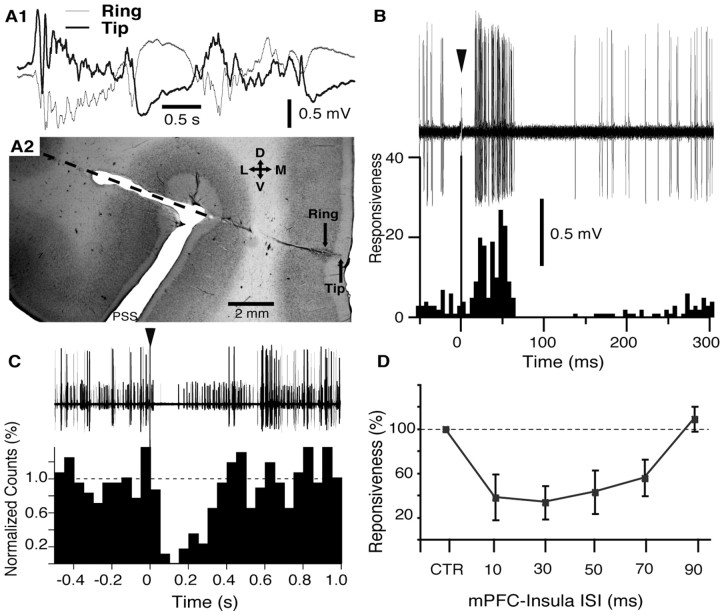
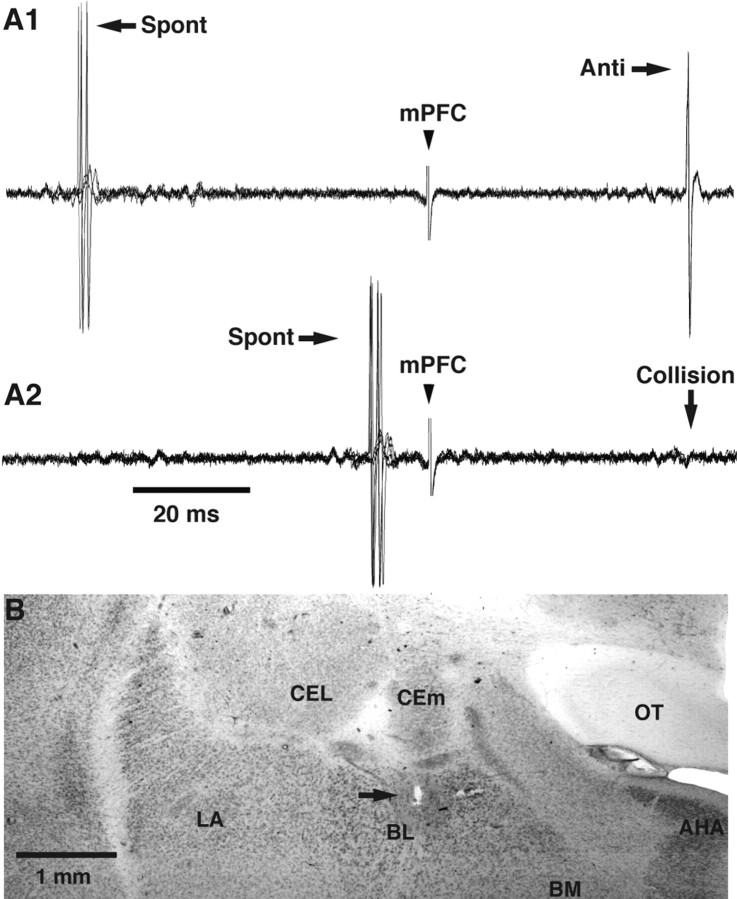
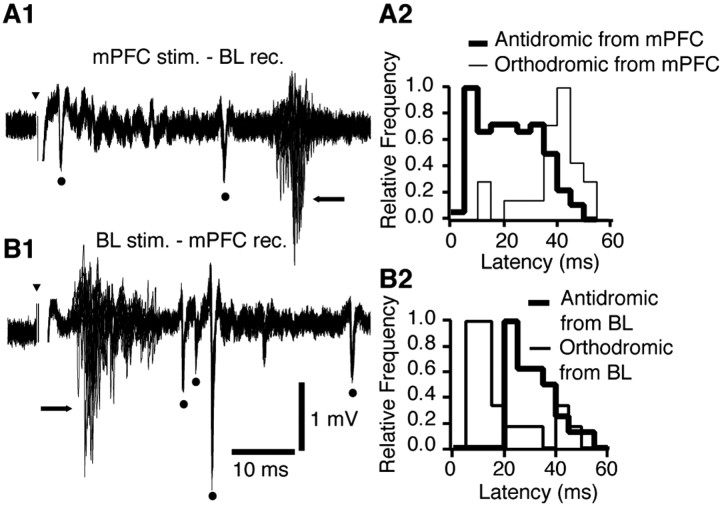
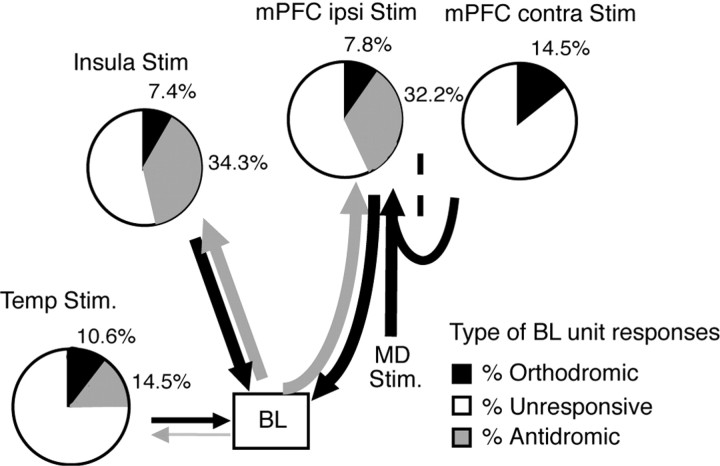
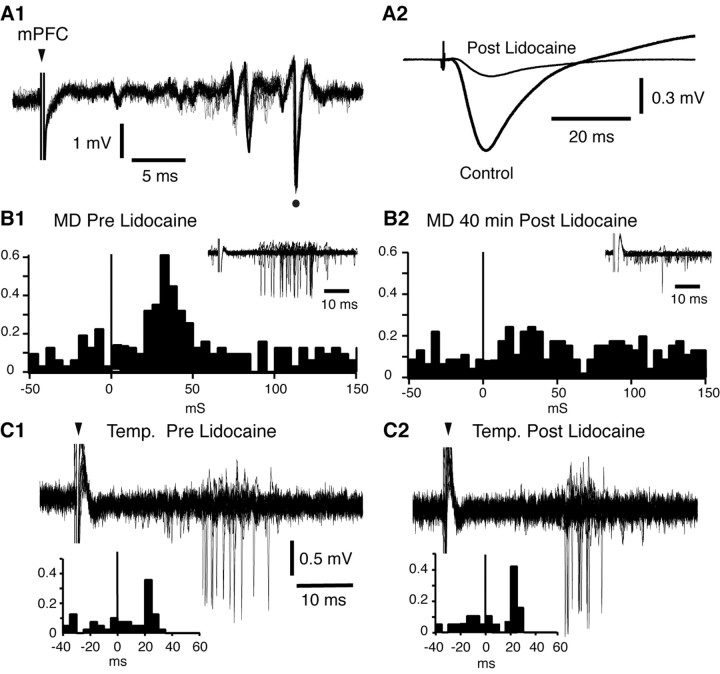
Accumulating evidence indicates that phobic and posttraumatic anxiety disorders likely result from a failure to extinguish fear memories. Extinction normally depends on a new learning that competes with the original fear memory and is driven by medial prefrontal cortex (mPFC) projections to the amygdala. Although mPFC stimulation was reported to inhibit the central medial (CEm) amygdala neurons that mediate fear responses via their brainstem and hypothalamic projections, it is unclear how this inhibition is generated. Because the mPFC has very sparse projections to CEm output neurons, the mPFC-evoked inhibition of the CEm is likely indirect. Thus, this study tested whether it resulted from a feedforward inhibition of basolateral amygdala (BLA) neurons that normally relay sensory inputs to the CEm. However, our results indicate that mPFC inputs excite rather than inhibit BLA neurons, implying that the inhibition of CEm cells is mediated by an active gating mechanism downstream of the BLA.
Figures
References
-
- Bechara A, Tranel D, Damasio H, Adolphs R, Rockland C, Damasio AR (1995) Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans. Science 269: 1115-1118. - PubMed
-
- Boehnke SE, Rasmusson DD (2001) Time course and effective spread of lidocaine and tetrodotoxin delivered via microdialysis: an electrophysiological study in cerebral cortex. J Neurosci Methods 105: 133-141. - PubMed
-
- Brinley-Reed M, Mascagni F, McDonald AJ (1995) Synaptology of prefrontal projections to the basolateral amygdala: an electron microscopic study in the rat. Neurosci Lett 202: 45-48. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources