

Distinct fibroblast growth factor (FGF)/FGF receptor signaling pairs initiate diverse cellular responses in the oligodendrocyte lineage
- PMID: 16093398
- PMCID: PMC6725305
- DOI: 10.1523/JNEUROSCI.2120-05.2005
Distinct fibroblast growth factor (FGF)/FGF receptor signaling pairs initiate diverse cellular responses in the oligodendrocyte lineage
Abstract
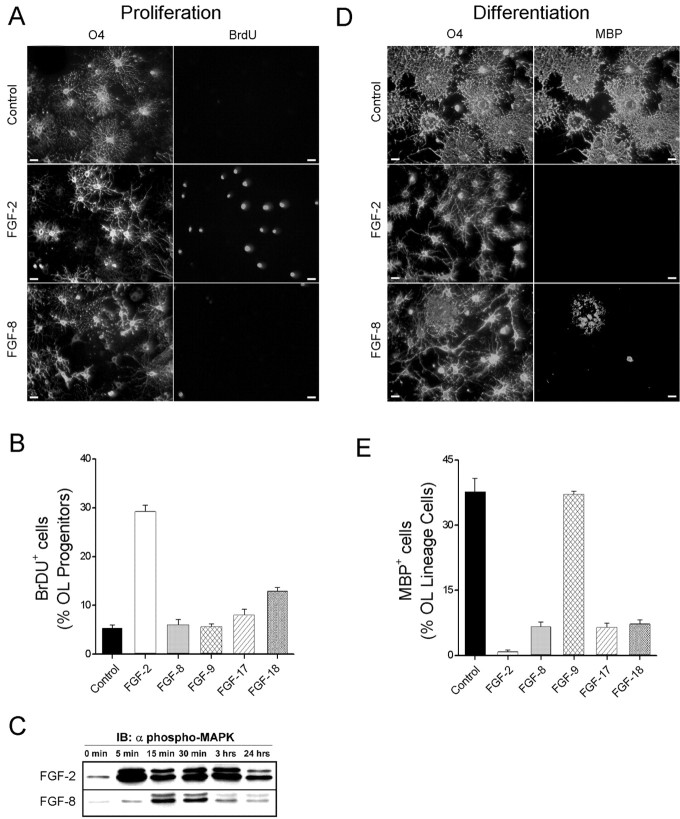
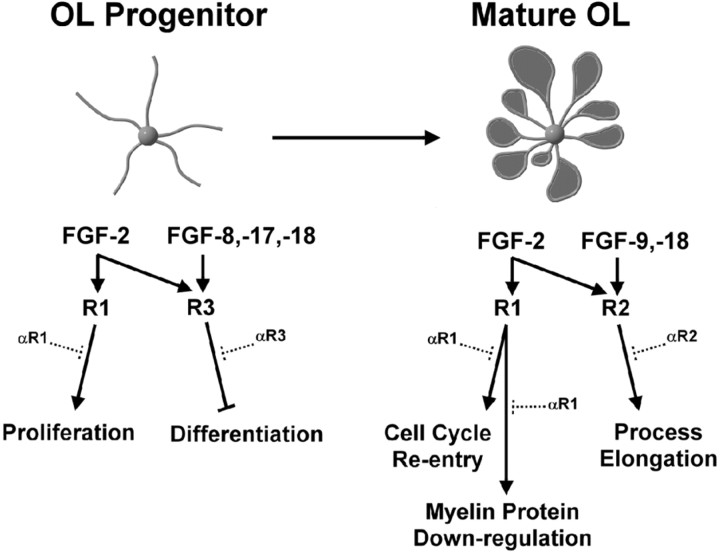
Fibroblast growth factors (FGFs) have been implicated in numerous cellular processes, including proliferation, migration, differentiation, and survival. Whereas FGF-2, the prototypic ligand in a family of 22 members, activates all four tyrosine kinase FGF receptors (FGFR1-FGFR4), other members demonstrate a higher degree of selectivity. Oligodendrocytes (OLs), the myelin-producing cells of the CNS, are highly influenced by FGF-2 at all stages of their development. However, how other FGFs and their cognate receptors orchestrate the development of OLs is essentially undefined. Using a combination of specific FGF ligands and receptor blocking antibodies, we now show that FGF-8 and FGF-17 target OL progenitors, inhibiting their terminal differentiation via the activation of FGFR3, whereas FGF-9 specifically targets differentiated OLs, triggering increases in process growth via FGFR2 signaling; FGF-18 targets both OL progenitors and OLs via activation of both FGFR2 and FGFR3. These events are highly correlated with changes in FGF receptor expression from FGFR3 to FGFR2 as OL progenitors differentiate into mature OLs. In addition, we demonstrate that, although activation of FGFR1 by FGF-2 leads to proliferation of OL progenitors, it produces deleterious effects on differentiated OLs (i.e., aberrant reentry into cell cycle and down-regulation of myelin proteins with a loss of myelin membrane). These data suggest that ligand availability, coupled with changes in FGF receptor expression, yield a changing repertoire of ligand-receptor signaling complexes that contribute critically to the regulation of both normal OL development and potential OL/myelin pathogenesis.
Figures






Similar articles
-
Phosphorylation and lipid raft association of fibroblast growth factor receptor-2 in oligodendrocytes.Glia. 2009 Jul;57(9):935-46. doi: 10.1002/glia.20818. Glia. 2009. PMID: 19053057 Free PMC article.
-
Specific inhibitor of FGF receptor signaling: FGF-2-mediated effects on proliferation, differentiation, and MAPK activation are inhibited by PD173074 in oligodendrocyte-lineage cells.J Neurosci Res. 2003 Nov 15;74(4):486-93. doi: 10.1002/jnr.10773. J Neurosci Res. 2003. PMID: 14598292
-
Differences in signal transduction pathways by which platelet-derived and fibroblast growth factors activate extracellular signal-regulated kinase in differentiating oligodendrocytes.J Neurochem. 2001 Mar;76(6):1925-34. doi: 10.1046/j.1471-4159.2001.00199.x. J Neurochem. 2001. PMID: 11259511
-
FGF-2 converts mature oligodendrocytes to a novel phenotype.J Neurosci Res. 1997 Oct 15;50(2):215-28. doi: 10.1002/(SICI)1097-4547(19971015)50:2<215::AID-JNR10>3.0.CO;2-7. J Neurosci Res. 1997. PMID: 9373031 Review.
-
Exploring mechanisms of FGF signalling through the lens of structural biology.Nat Rev Mol Cell Biol. 2013 Mar;14(3):166-80. doi: 10.1038/nrm3528. Epub 2013 Feb 13. Nat Rev Mol Cell Biol. 2013. PMID: 23403721 Free PMC article. Review.
Cited by
-
Toxicarioside A inhibits SGC-7901 proliferation, migration and invasion via NF-κB/bFGF signaling.World J Gastroenterol. 2012 Apr 14;18(14):1602-9. doi: 10.3748/wjg.v18.i14.1602. World J Gastroenterol. 2012. PMID: 22529688 Free PMC article.
-
Multiple Sclerosis and Aging: The Dynamics of Demyelination and Remyelination.ASN Neuro. 2022 Jan-Dec;14:17590914221118502. doi: 10.1177/17590914221118502. ASN Neuro. 2022. PMID: 35938615 Free PMC article. Review.
-
Monoclonal antibodies to fibroblast growth factor receptor 2 effectively inhibit growth of gastric tumor xenografts.Clin Cancer Res. 2010 Dec 1;16(23):5750-8. doi: 10.1158/1078-0432.CCR-10-0531. Epub 2010 Jul 29. Clin Cancer Res. 2010. PMID: 20670946 Free PMC article.
-
Quantitative and qualitative differences in the activation of a fibroblast growth factor receptor by different FGF ligands.Cytokine Growth Factor Rev. 2024 Aug;78:77-84. doi: 10.1016/j.cytogfr.2024.07.002. Epub 2024 Jul 9. Cytokine Growth Factor Rev. 2024. PMID: 39043538 Free PMC article. Review.
-
Exogenous FGF-1 Differently Regulates Oligodendrocyte Replenishment in an SCI Repair Model and Cultured Cells.Biomedicines. 2022 Oct 27;10(11):2724. doi: 10.3390/biomedicines10112724. Biomedicines. 2022. PMID: 36359244 Free PMC article.
References
-
- Asai T, Wanaka A, Kato H, Masana Y, Seo M, Tohyama M (1993) Differential expression of two members of FGF receptor gene family, FGFR-1 and FGFR-2 mRNA, in the adult rat central nervous system. Mol Brain Res 17: 174-178. - PubMed
-
- Bansal R (2002) Fibroblast growth factors and their receptors in oligodendrocyte development: implications for demyelination and remyelination. Dev Neurosci 46: 24-35. - PubMed
-
- Bansal R, Pfeiffer SE (1994) Inhibition of protein and lipid sulfation in oligodendrocytes blocks biological responses to FGF-2 and retards cytoarchitectural maturation, but not developmental lineage progression. Dev Biol 162: 511-524. - PubMed
-
- Bansal R, Pfeiffer SE (1997) FGF-2 converts mature oligodendrocytes to a novel phenotype. J Neurosci Res 50: 215-228. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous