

The E-hook of tubulin interacts with kinesin's head to increase processivity and speed
- PMID: 16100283
- PMCID: PMC1366818
- DOI: 10.1529/biophysj.104.057505
The E-hook of tubulin interacts with kinesin's head to increase processivity and speed
Abstract
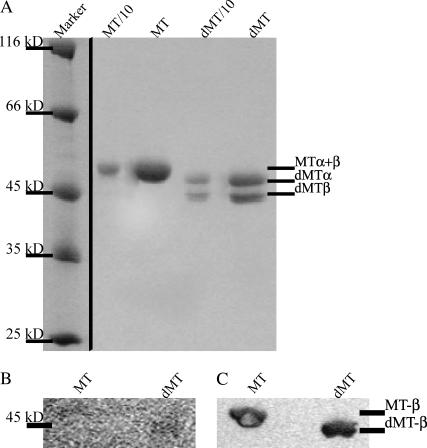
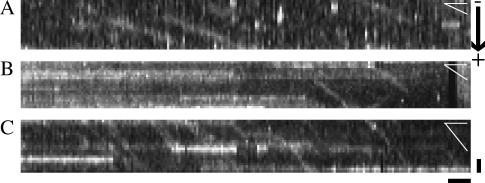
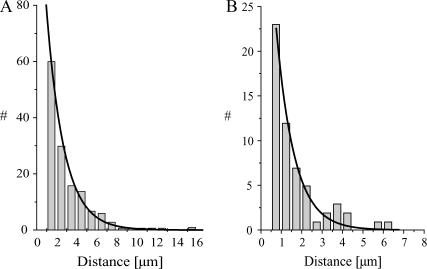
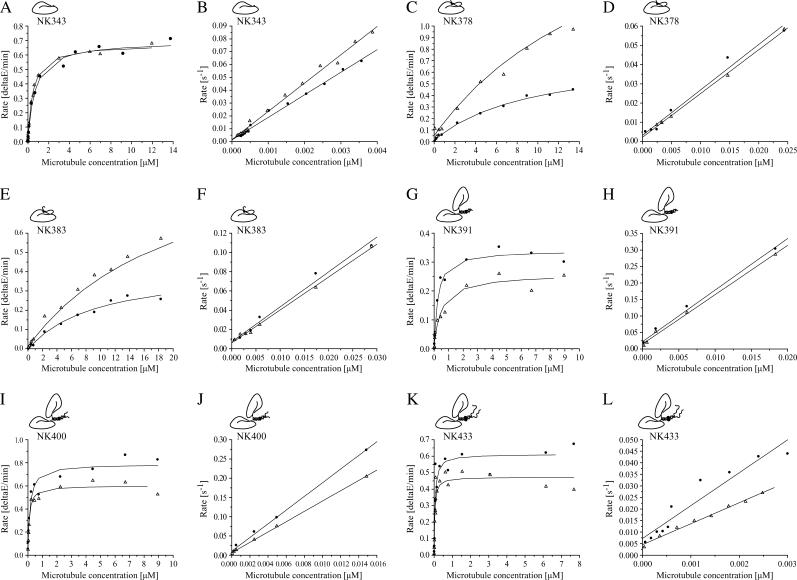
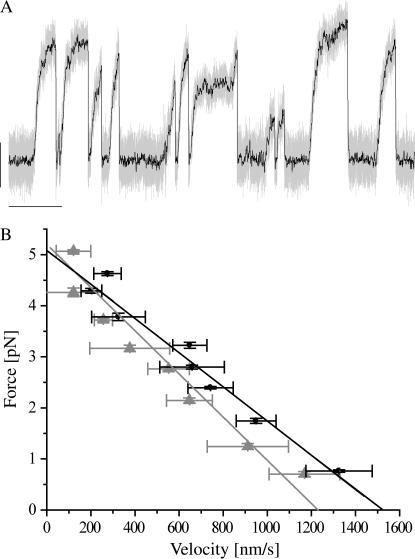
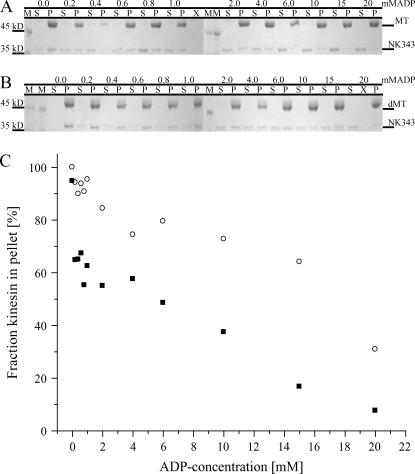
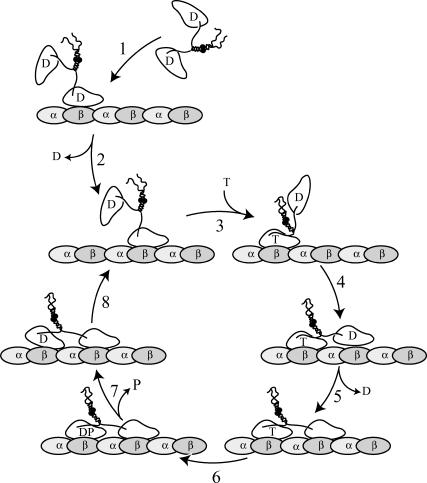
Kinesins are dimeric motor proteins that move processively along microtubules. It has been proposed that the processivity of conventional kinesins is increased by electrostatic interactions between the positively charged neck of the motor and the negatively charged C-terminus of tubulin (E-hook). In this report we challenge this anchoring hypothesis by studying the motility of a fast fungal kinesin from Neurospora crassa (NcKin). NcKin is highly processive despite lacking the positive charges in the neck. We present a detailed analysis of how proteolytic removal of the E-hook affects truncated monomeric and dimeric constructs of NcKin. Upon digestion we observe a strong reduction of the processivity and speed of dimeric motor constructs. Monomeric motors with truncated or no neck display the same reduction of microtubule gliding speed as dimeric constructs, suggesting that the E-hook interacts with the head only. The E-hook has no effect on the strongly bound states of NcKin as microtubule digestion does not alter the stall forces produced by single dimeric motors, suggesting that the E-hook affects the interaction site of the kinesin.ADP-head and the microtubule. In fact, kinetic and binding experiments indicate that removal of the E-hook shifts the binding equilibrium of the weakly attached kinesin.ADP-head toward a more strongly bound state, which may explain reduced processivity and speed on digested microtubules.
Figures
Similar articles
-
Unusual properties of the fungal conventional kinesin neck domain from Neurospora crassa.EMBO J. 2001 Nov 15;20(22):6226-35. doi: 10.1093/emboj/20.22.6226. EMBO J. 2001. PMID: 11707394 Free PMC article.
-
A new look at the microtubule binding patterns of dimeric kinesins.J Mol Biol. 2000 Apr 14;297(5):1087-103. doi: 10.1006/jmbi.2000.3627. J Mol Biol. 2000. PMID: 10764575
-
Structure of a fast kinesin: implications for ATPase mechanism and interactions with microtubules.EMBO J. 2001 Nov 15;20(22):6213-25. doi: 10.1093/emboj/20.22.6213. EMBO J. 2001. PMID: 11707393 Free PMC article.
-
Kinesin, 30 years later: Recent insights from structural studies.Protein Sci. 2015 Jul;24(7):1047-56. doi: 10.1002/pro.2697. Epub 2015 Jun 11. Protein Sci. 2015. PMID: 25975756 Free PMC article. Review.
-
These motors were made for walking.Protein Sci. 2020 Aug;29(8):1707-1723. doi: 10.1002/pro.3895. Epub 2020 Jun 26. Protein Sci. 2020. PMID: 32472639 Free PMC article. Review.
Cited by
-
Computational Study on E-Hooks of Tubulins in the Binding Process with Kinesin.Int J Mol Sci. 2022 Feb 12;23(4):2035. doi: 10.3390/ijms23042035. Int J Mol Sci. 2022. PMID: 35216151 Free PMC article.
-
Myosin V and Kinesin act as tethers to enhance each others' processivity.Proc Natl Acad Sci U S A. 2008 Mar 25;105(12):4691-6. doi: 10.1073/pnas.0711531105. Epub 2008 Mar 17. Proc Natl Acad Sci U S A. 2008. PMID: 18347333 Free PMC article.
-
The motility of axonemal dynein is regulated by the tubulin code.Biophys J. 2014 Dec 16;107(12):2872-2880. doi: 10.1016/j.bpj.2014.10.061. Biophys J. 2014. PMID: 25658008 Free PMC article.
-
A chimeric kinesin-1 head/kinesin-5 tail motor switches between diffusive and processive motility.Biophys J. 2013 Jan 22;104(2):432-41. doi: 10.1016/j.bpj.2012.11.3810. Biophys J. 2013. PMID: 23442865 Free PMC article.
-
Collective Force Regulation in Anti-parallel Microtubule Gliding by Dimeric Kif15 Kinesin Motors.Curr Biol. 2017 Sep 25;27(18):2810-2820.e6. doi: 10.1016/j.cub.2017.08.018. Epub 2017 Sep 14. Curr Biol. 2017. PMID: 28918951 Free PMC article.
References
-
- Vale, R. D. 2003. The molecular motor toolbox for intracellular transport. Cell. 112:467–480. - PubMed
-
- Brady, S. T. 1985. A novel brain ATPase with properties expected for the fast axonal transport motor. Nature. 317:73–75. - PubMed
-
- Scholey, J. M., J. Heuser, J. T. Yang, and L. S. Goldstein. 1989. Identification of globular mechanochemical heads of kinesin. Nature. 338:355–357. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
