

CD46-utilizing adenoviruses inhibit C/EBPbeta-dependent expression of proinflammatory cytokines
- PMID: 16103178
- PMCID: PMC1193609
- DOI: 10.1128/JVI.79.17.11259-11268.2005
CD46-utilizing adenoviruses inhibit C/EBPbeta-dependent expression of proinflammatory cytokines
Abstract
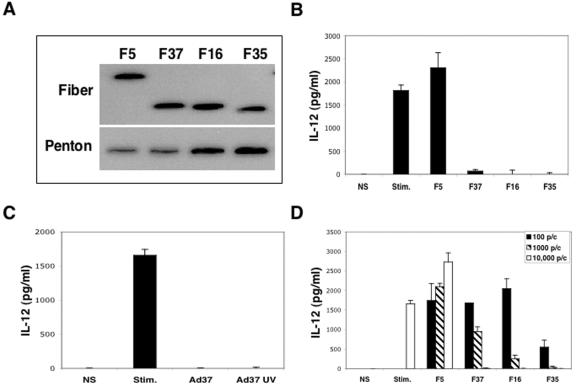
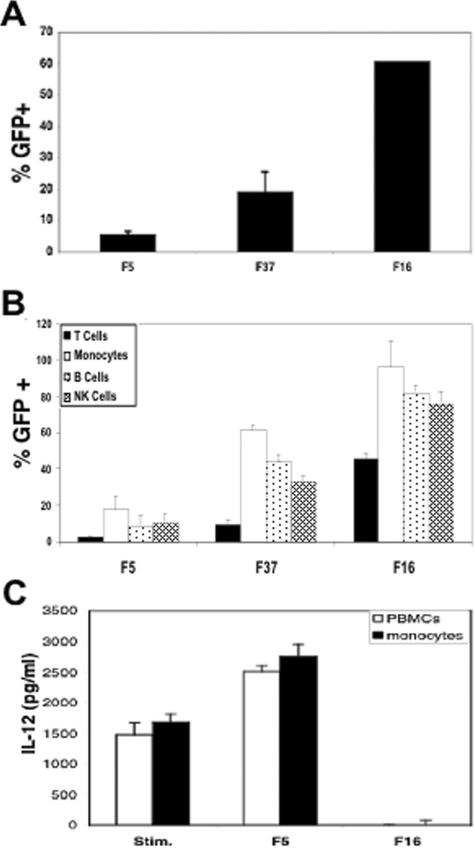
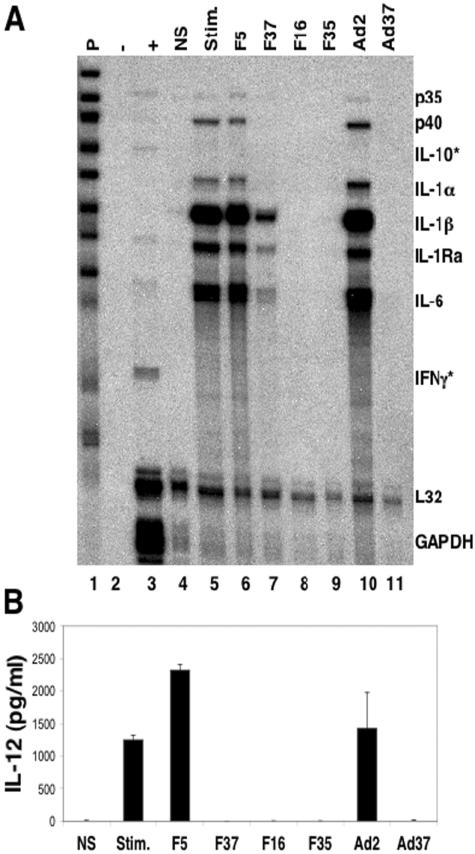
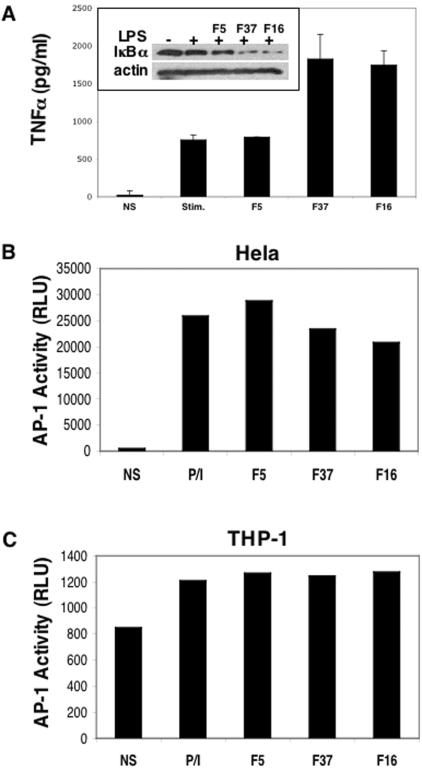
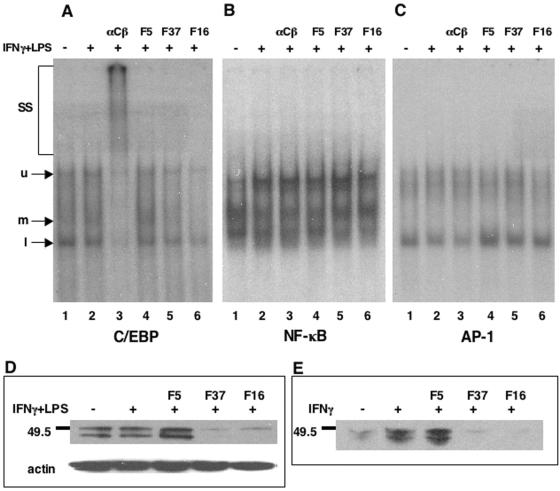
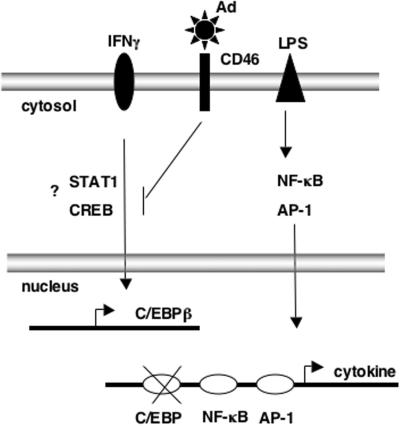
The majority of adenovirus serotypes utilize the coxsackievirus-adenovirus receptor (CAR) for virus-host cell attachment, but subgroup B and subgroup D (adenovirus type 37 [Ad37]) viruses recognize CD46. CD46 is a ubiquitously expressed receptor that serves as a cofactor for the inactivation of the complement components C3b and C4b, and it also serves as a receptor for diverse microbial pathogens. A reported consequence of CD46 engagement is a reduced capability of human immune cells to express interleukin-12 (IL-12), a cytokine involved in both the innate and adaptive immune responses. Studies were thus undertaken to determine whether CD46-utilizing Ads alter the expression of proinflammatory cytokines. Subgroup B (Ad16 and -35) and Ad37, but not Ad2 or -5, significantly reduced IL-12 production by human peripheral blood mononuclear cells stimulated with gamma interferon (IFN-gamma) and lipopolysaccharide. IL-12 mRNA (p35 and p40 subunits) levels as well as other cytokine mRNA levels (IL-1alpha and -beta, IL-1Ra, and IL-6) were decreased upon interaction with CD46-utilizing Ads. Analysis of transcription factor activity required for cytokine expression indicated that CD46-utilizing Ads preferentially inhibited IFN-gamma-induced C/EBPbeta protein expression, consequently reducing its ability to form DNA complexes. Interference with IFN-gamma signaling events by CD46-utilizing Ads, but not CAR-utilizing Ads, reveals a potentially critical difference in the host immune response against distinct Ad vectors, a situation that has implications for gene delivery and vaccine development.
Figures
References
-
- Astier, A., M. C. Trescol-Biemont, O. Azocar, B. Lamouille, and C. Rabourdin-Combe. 2000. Cutting edge: CD46, a new costimulatory molecule for T cells, that induces p120CBL and LAT phosphorylation. J. Immunol. 164:6091-6095. - PubMed
-
- Atabani, S. F., A. A. Byrnes, A. Jaye, I. M. Kidd, A. F. Magnusen, H. Whittle, and C. L. Karp. 2001. Natural measles causes prolonged suppression of interleukin-12 production. J. Infect. Dis. 184:1-9. - PubMed
-
- Awad, S., H. Yokozeki, Y. Miyazaki, K. Igawa, K. Minatohara, T. Satoh, and K. Nishioka. 2002. Glucocorticoids induced the production and gene expression of IL-1α through AP-1 and partially NF-κB activation in murine epidermal cells. J. Med. Dent. Sci. 49:27-35. - PubMed
-
- Baccam, M., S. Y. Woo, C. Vinson, and G. A. Bishop. 2003. CD40-mediated transcriptional regulation of the IL-6 gene in B lymphocytes: involvement of NF-κB, AP-1, and C/EBP. J. Immunol. 170:3099-3108. - PubMed
-
- Becker, C., S. Wirtz, X. Ma, M. Blessing, P. R. Galle, and M. F. Neurath. 2001. Regulation of IL-12 p40 promoter activity in primary human monocytes: roles of NF-κB, CCAAT/enhancer-binding protein β, and PU.1 and identification of a novel repressor element (GA-12) that responds to IL-4 and prostaglandin E2. J. Immunol. 167:2608-2618. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
