

Cyclic AMP and calcium interplay as second messengers in melatonin-dependent regulation of Plasmodium falciparum cell cycle
- PMID: 16103224
- PMCID: PMC2171486
- DOI: 10.1083/jcb.200505117
Cyclic AMP and calcium interplay as second messengers in melatonin-dependent regulation of Plasmodium falciparum cell cycle
Abstract
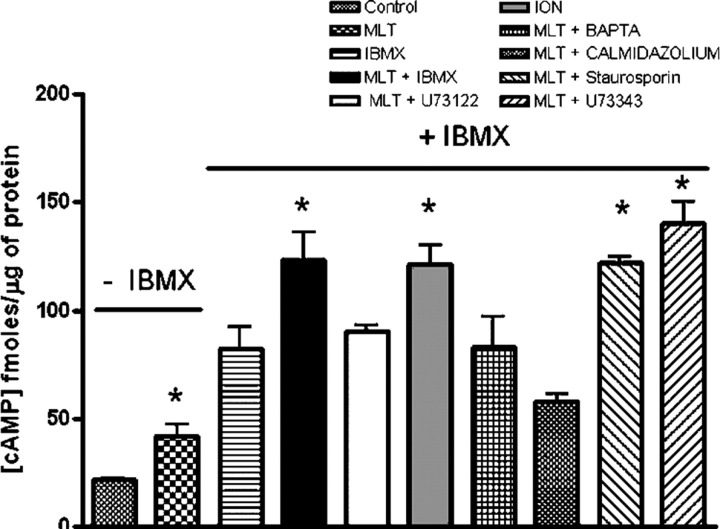
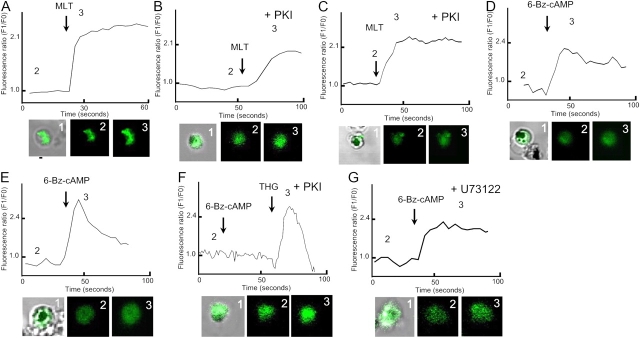
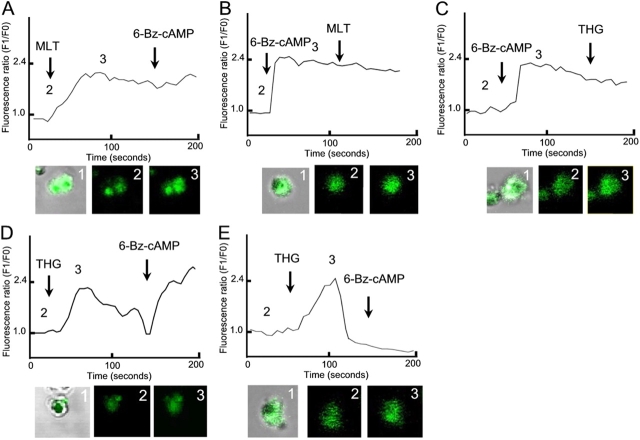
The host hormone melatonin increases cytoplasmic Ca(2+) concentration and synchronizes Plasmodium cell cycle (Hotta, C.T., M.L. Gazarini, F.H. Beraldo, F.P. Varotti, C. Lopes, R.P. Markus, T. Pozzan, and C.R. Garcia. 2000. Nat. Cell Biol. 2:466-468). Here we show that in Plasmodium falciparum melatonin induces an increase in cyclic AMP (cAMP) levels and cAMP-dependent protein kinase (PKA) activity (40 and 50%, respectively). When red blood cells infected with P. falciparum are treated with cAMP analogue adenosine 3',5'-cyclic monophosphate N6-benzoyl/PKA activator (6-Bz-cAMP) there is an alteration of the parasite cell cycle. This effect appears to depend on activation of PKA (abolished by the PKA inhibitors adenosine 3',5'-cyclic monophosphorothioate/8 Bromo Rp isomer, PKI [cell permeable peptide], and H89). An unexpected cross talk was found to exist between the cAMP and the Ca(2+)-dependent signaling pathways. The increases in cAMP by melatonin are inhibited by blocker of phospholipase C U73122, and addition of 6-Bz-cAMP increases cytosolic Ca(2+) concentration, through PKA activation. These findings suggest that in Plasmodium a highly complex interplay exists between the Ca(2+) and cAMP signaling pathways, but also that the control of the parasite cell cycle by melatonin requires the activation of both second messenger controlled pathways.
Figures



Similar articles
-
Melatonin triggers PKA activation in the rodent malaria parasite Plasmodium chabaudi.J Pineal Res. 2011 Jan;50(1):64-70. doi: 10.1111/j.1600-079X.2010.00810.x. Epub 2010 Oct 22. J Pineal Res. 2011. PMID: 20964707
-
An interplay between 2 signaling pathways: melatonin-cAMP and IP3-Ca2+ signaling pathways control intraerythrocytic development of the malaria parasite Plasmodium falciparum.Biochem Biophys Res Commun. 2014 Mar 28;446(1):125-31. doi: 10.1016/j.bbrc.2014.02.070. Epub 2014 Mar 4. Biochem Biophys Res Commun. 2014. PMID: 24607908
-
Elevated extracellular calcium increases fibroblast growth factor-2 gene and protein expression levels via a cAMP/PKA dependent pathway in cementoblasts.Bone. 2010 Sep;47(3):564-72. doi: 10.1016/j.bone.2010.05.042. Epub 2010 Jun 9. Bone. 2010. PMID: 20542497
-
cAMP-dependent protein kinase from Plasmodium falciparum: an update.Parasitology. 2011 Jan;138(1):1-25. doi: 10.1017/S003118201000096X. Epub 2010 Jul 21. Parasitology. 2011. PMID: 20663247 Review.
-
PKA and Apicomplexan Parasite Diseases.Horm Metab Res. 2017 Apr;49(4):296-300. doi: 10.1055/s-0042-118459. Epub 2016 Nov 11. Horm Metab Res. 2017. PMID: 27835919 Free PMC article. Review.
Cited by
-
Noggin is novel inducer of mesenchymal stem cell adipogenesis: implications for bone health and obesity.J Biol Chem. 2012 Apr 6;287(15):12241-9. doi: 10.1074/jbc.M111.293613. Epub 2012 Feb 20. J Biol Chem. 2012. PMID: 22351751 Free PMC article.
-
Blockage of spontaneous Ca2+ oscillation causes cell death in intraerythrocitic Plasmodium falciparum.PLoS One. 2012;7(7):e39499. doi: 10.1371/journal.pone.0039499. Epub 2012 Jul 6. PLoS One. 2012. PMID: 22792177 Free PMC article.
-
Phenotypic and transcriptomic analyses of Plasmodium falciparum protein kinase A catalytic subunit inhibition.Parasitol Res. 2009 Nov;105(6):1691-9. doi: 10.1007/s00436-009-1615-6. Epub 2009 Sep 25. Parasitol Res. 2009. PMID: 19779742
-
Malaria parasites and circadian rhythm: New insights into an old puzzle.Curr Res Microb Sci. 2020 Dec 14;2:100017. doi: 10.1016/j.crmicr.2020.100017. eCollection 2021 Dec. Curr Res Microb Sci. 2020. PMID: 34841309 Free PMC article.
-
Melatonin and IP3-induced Ca2+ release from intracellular stores in the malaria parasite Plasmodium falciparum within infected red blood cells.J Biol Chem. 2011 Feb 18;286(7):5905-12. doi: 10.1074/jbc.M110.188474. Epub 2010 Dec 13. J Biol Chem. 2011. PMID: 21149448 Free PMC article.
References
-
- Alleva, L.M., and K. Kirk. 2001. Calcium regulation in the intraerythrocytic malaria parasite Plasmodium falciparum. Mol. Biochem. Parasitol. 117:121–128. - PubMed
-
- Aravind L., L.M. Iyer, T.E. Wellems, and L.H. Miller. 2003. Plasmodium biology: genomic gleanings. Cell. 115:771–785. - PubMed
-
- Bannister, L., and G. Mitchell. 2003. The ins, outs and roundabouts of malaria. Trends Parasitol. 19:209–213. - PubMed
-
- Beraldo, F.H., and C.R. Garcia. 2005. Products of tryptophan catabolism induce a Ca2+ release and modulate the cell cycle of P. fal ciparum malaria parasites J. Pineal Res. In press. - PubMed
-
- Berridge, M.J., M.D. Bootman, and H.L. Roderick. 2003. Calcium signalling: dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 4:517–529. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
