

Inducible DNA breaks in Ig S regions are dependent on AID and UNG
- PMID: 16103411
- PMCID: PMC2212854
- DOI: 10.1084/jem.20050872
Inducible DNA breaks in Ig S regions are dependent on AID and UNG
Abstract
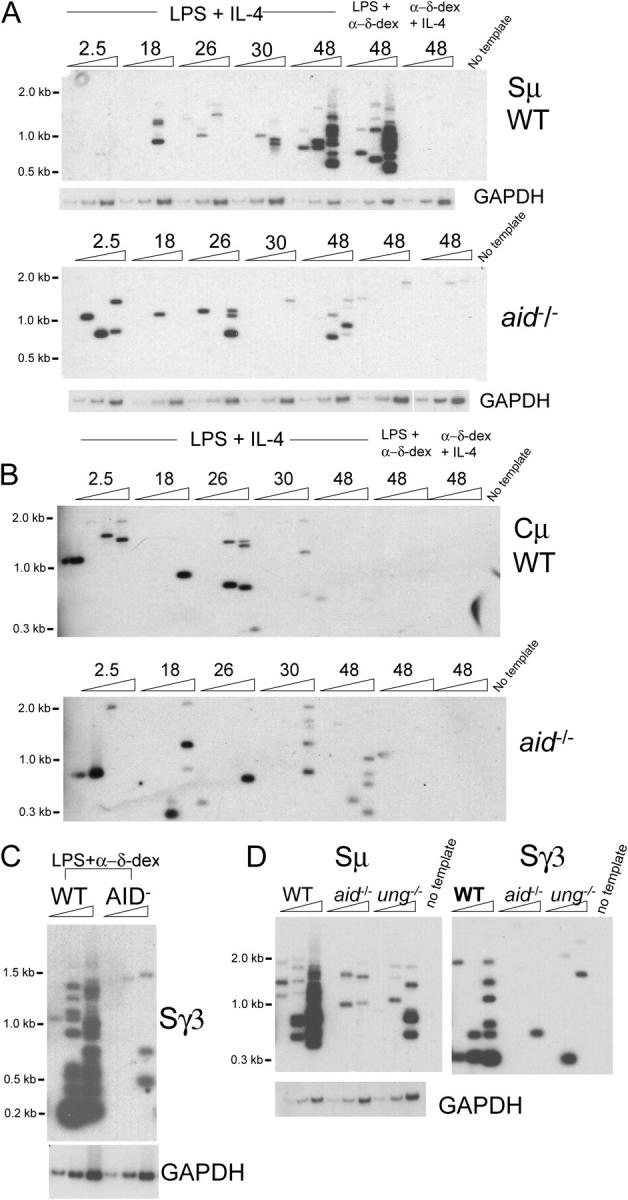
Class switch recombination (CSR) occurs by an intrachromosomal deletion whereby the IgM constant region gene (Cmu) is replaced by a downstream constant region gene. This unique recombination event involves formation of double-strand breaks (DSBs) in immunoglobulin switch (S) regions, and requires activation-induced cytidine deaminase (AID), which converts cytosines to uracils. Repair of the uracils is proposed to lead to DNA breaks required for recombination. Uracil DNA glycosylase (UNG) is required for most CSR activity although its role is disputed. Here we use ligation-mediated PCR to detect DSBs in S regions in splenic B cells undergoing CSR. We find that the kinetics of DSB induction corresponds with AID expression, and that DSBs are AID- and UNG-dependent and occur preferentially at G:C basepairs in WRC/GYW AID hotspots. Our results indicate that AID attacks cytosines on both DNA strands, and staggered breaks are processed to blunt DSBs at the initiating ss break sites. We propose a model to explain the types of end-processing events observed.
Figures



Similar articles
-
Single-strand DNA breaks in Ig class switch recombination that depend on UNG but not AID.Int Immunol. 2008 Nov;20(11):1381-93. doi: 10.1093/intimm/dxn097. Epub 2008 Sep 15. Int Immunol. 2008. PMID: 18794203
-
AID- and Ung-dependent generation of staggered double-strand DNA breaks in immunoglobulin class switch DNA recombination: a post-cleavage role for AID.Mol Immunol. 2008 Nov;46(1):45-61. doi: 10.1016/j.molimm.2008.07.003. Epub 2008 Aug 28. Mol Immunol. 2008. PMID: 18760480 Free PMC article.
-
Non-canonical uracil processing in DNA gives rise to double-strand breaks and deletions: relevance to class switch recombination.Nucleic Acids Res. 2016 Apr 7;44(6):2691-705. doi: 10.1093/nar/gkv1535. Epub 2016 Jan 6. Nucleic Acids Res. 2016. PMID: 26743004 Free PMC article.
-
The contested role of uracil DNA glycosylase in immunoglobulin gene diversification.Trends Genet. 2005 May;21(5):253-6. doi: 10.1016/j.tig.2005.02.013. Trends Genet. 2005. PMID: 15851057 Review.
-
Pathophysiology of B-cell intrinsic immunoglobulin class switch recombination deficiencies.Adv Immunol. 2007;94:275-306. doi: 10.1016/S0065-2776(06)94009-7. Adv Immunol. 2007. PMID: 17560278 Review.
Cited by
-
APOBEC3G protects the genome of human cultured cells and mice from radiation-induced damage.FEBS J. 2023 Apr;290(7):1822-1839. doi: 10.1111/febs.16673. Epub 2022 Nov 25. FEBS J. 2023. PMID: 36325681 Free PMC article.
-
Activation induced cytidine deaminase: An old friend with new faces.Front Immunol. 2022 Oct 27;13:965312. doi: 10.3389/fimmu.2022.965312. eCollection 2022. Front Immunol. 2022. PMID: 36405752 Free PMC article. Review.
-
Tracking Immunoglobulin Repertoire and Transcriptomic Changes in Germinal Center B Cells by Single-Cell Analysis.Front Immunol. 2022 Jan 12;12:818758. doi: 10.3389/fimmu.2021.818758. eCollection 2021. Front Immunol. 2022. PMID: 35095922 Free PMC article.
-
AID recruits UNG and Msh2 to Ig switch regions dependent upon the AID C terminus [corrected].J Immunol. 2011 Sep 1;187(5):2464-75. doi: 10.4049/jimmunol.1101406. Epub 2011 Jul 29. J Immunol. 2011. PMID: 21804017 Free PMC article.
-
SAMHD1-mediated dNTP degradation is required for efficient DNA repair during antibody class switch recombination.EMBO J. 2020 Aug 3;39(15):e102931. doi: 10.15252/embj.2019102931. Epub 2020 Jun 8. EMBO J. 2020. PMID: 32511795 Free PMC article.
References
-
- Stavnezer, J. 2000. Molecular processes that regulate class switching. Curr. Top. Microbiol. Immunol. 245:127–168. - PubMed
-
- Manis, J.P., M. Tian, and F.W. Alt. 2002. Mechanism and control of class-switch recombination. Trends Immunol. 23:31–39. - PubMed
-
- Muramatsu, M., K. Kinoshita, S. Fagarasan, S. Yamada, Y. Shinkai, and T. Honjo. 2000. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 102:553–563. - PubMed
-
- Revy, P., T. Muto, Y. Levy, F. Geissmann, A. Plebani, O. Sanal, N. Catalan, M. Forveille, R. Dufourcq-Labelouse, A. Gennery, et al. 2000. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2). Cell. 102:565–575. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
