

Pseudogenization of a sweet-receptor gene accounts for cats' indifference toward sugar
- PMID: 16103917
- PMCID: PMC1183522
- DOI: 10.1371/journal.pgen.0010003
Pseudogenization of a sweet-receptor gene accounts for cats' indifference toward sugar
Abstract
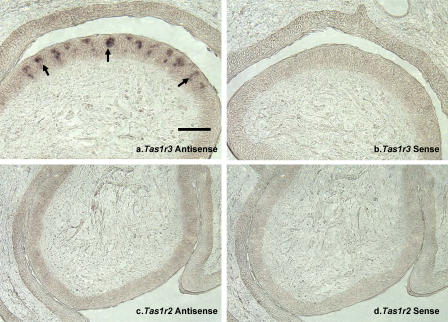
Although domestic cats (Felis silvestris catus) possess an otherwise functional sense of taste, they, unlike most mammals, do not prefer and may be unable to detect the sweetness of sugars. One possible explanation for this behavior is that cats lack the sensory system to taste sugars and therefore are indifferent to them. Drawing on work in mice, demonstrating that alleles of sweet-receptor genes predict low sugar intake, we examined the possibility that genes involved in the initial transduction of sweet perception might account for the indifference to sweet-tasting foods by cats. We characterized the sweet-receptor genes of domestic cats as well as those of other members of the Felidae family of obligate carnivores, tiger and cheetah. Because the mammalian sweet-taste receptor is formed by the dimerization of two proteins (T1R2 and T1R3; gene symbols Tas1r2 and Tas1r3), we identified and sequenced both genes in the cat by screening a feline genomic BAC library and by performing PCR with degenerate primers on cat genomic DNA. Gene expression was assessed by RT-PCR of taste tissue, in situ hybridization, and immunohistochemistry. The cat Tas1r3 gene shows high sequence similarity with functional Tas1r3 genes of other species. Message from Tas1r3 was detected by RT-PCR of taste tissue. In situ hybridization and immunohistochemical studies demonstrate that Tas1r3 is expressed, as expected, in taste buds. However, the cat Tas1r2 gene shows a 247-base pair microdeletion in exon 3 and stop codons in exons 4 and 6. There was no evidence of detectable mRNA from cat Tas1r2 by RT-PCR or in situ hybridization, and no evidence of protein expression by immunohistochemistry. Tas1r2 in tiger and cheetah and in six healthy adult domestic cats all show the similar deletion and stop codons. We conclude that cat Tas1r3 is an apparently functional and expressed receptor but that cat Tas1r2 is an unexpressed pseudogene. A functional sweet-taste receptor heteromer cannot form, and thus the cat lacks the receptor likely necessary for detection of sweet stimuli. This molecular change was very likely an important event in the evolution of the cat's carnivorous behavior.
Conflict of interest statement
Competing interests. HW, JC, KM, and LH declare that they have no competing interests of a financial, professional, or personal nature. VLD is an employee of the Masterfoods division of Mars. GKB is on an advisory board to the WALTHAM Centre. Patents describing the uses of the feline receptors have been filed, and name as inventors: XL, WL, JGB, DRR, and AAB.
Figures



References
-
- Beauchamp GK, Maller O, Rogers JG. Flavor preferences in cats (Felis catus and Panthera sp.) J Comp Physiol Psychol. 1977;91:1118–1127.
-
- Carpenter JA. Species differences in taste preferences. J Comp Physiol Psychol. 1956;49:139–144. - PubMed
-
- Bartoshuk LM, Jacobs HL, Nichols TL, Hoff LA, Ryckman JJ. Taste rejection of nonnutritive sweeteners in cats. J Comp Physiol Psychol. 1975;89:971–975. - PubMed
-
- Bradshaw JW. Sensory and experiential factors in the design of foods for domestic dogs and cats. Proc Nutr Soc. 1991;50:99–106. - PubMed
-
- White TD, Boudreau JC. Taste preferences of the cat for neurophysiologically active compounds. Physiol Psychol. 1975;3:405–410.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
