

Strong effects of subphysiological temperature on the function and plasticity of mammalian presynaptic terminals
- PMID: 16107635
- PMCID: PMC6725406
- DOI: 10.1523/JNEUROSCI.1801-05.2005
Strong effects of subphysiological temperature on the function and plasticity of mammalian presynaptic terminals
Abstract
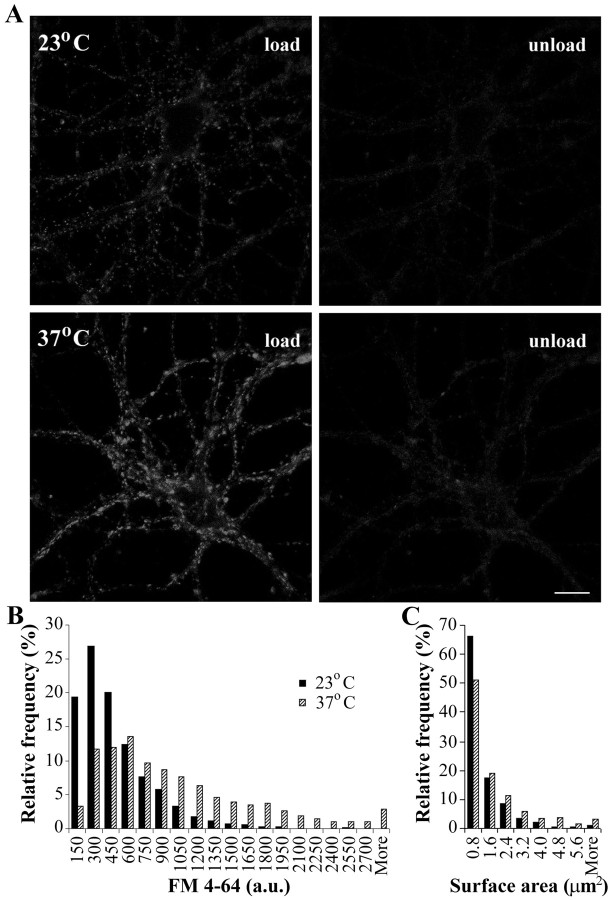
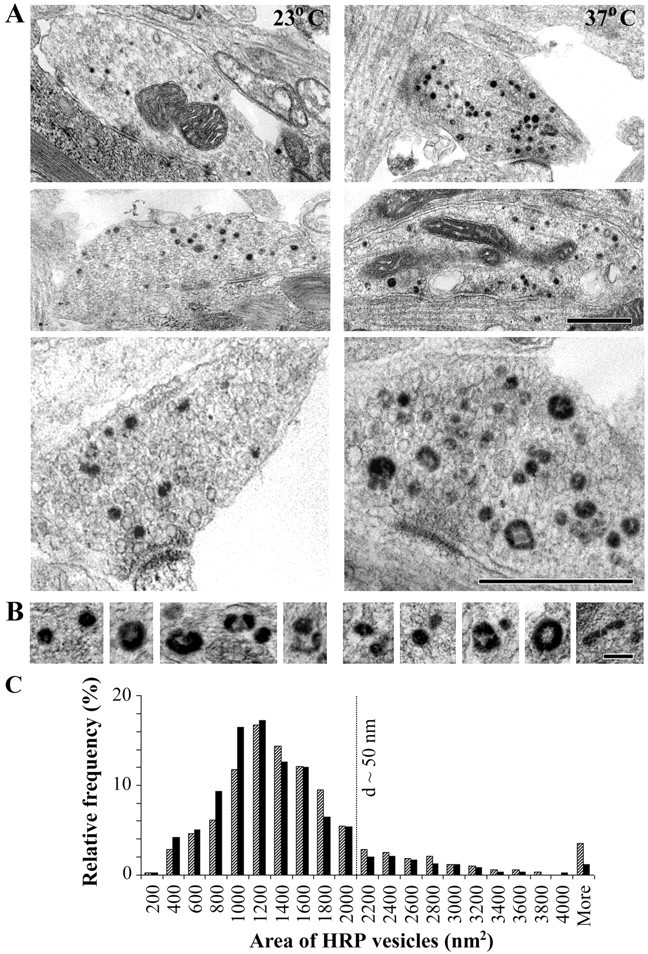
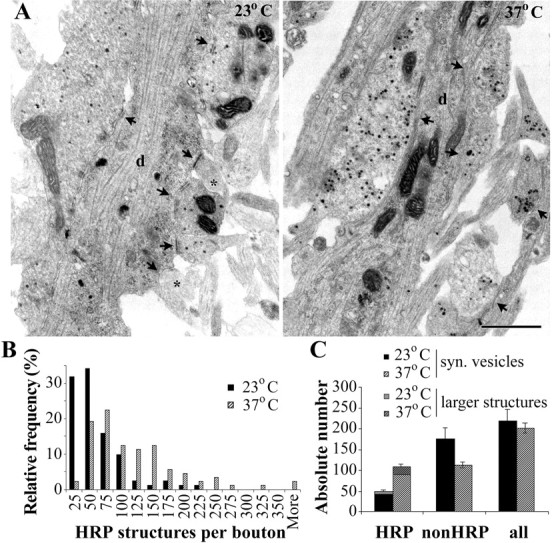
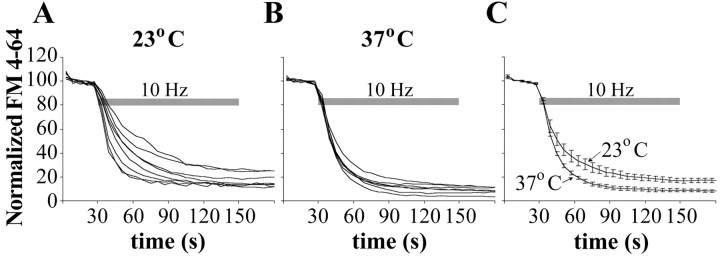
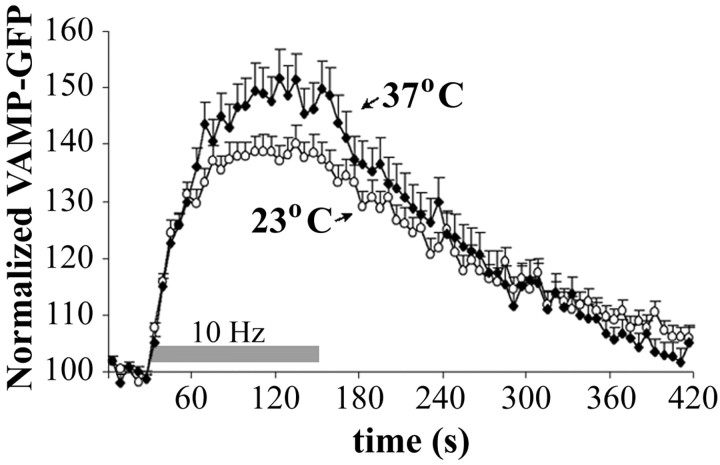
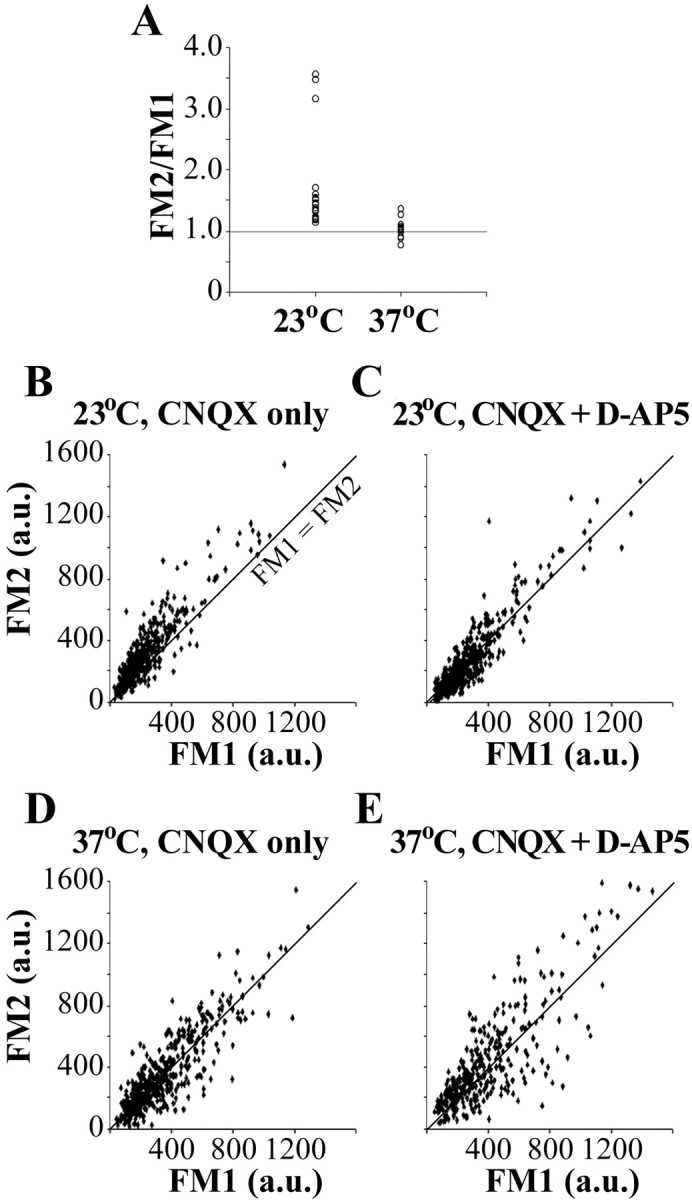
Most cellular processes are known to be strongly temperature dependent. Nevertheless, a large fraction of studies of mammalian synaptic function have been and are performed near room temperature (i.e., at least 10 degrees C below physiological temperature). Here, we examined the effects of temperature on presynaptic function in primary cultures of rat hippocampal neurons. FM dyes, VAMP (vesicle-associated membrane protein)-GFP (green fluorescent protein) transfection, and HRP uptake were used to quantify various aspects of synaptic vesicle recycling. Our results show that there are very substantial differences in synaptic vesicle recycling at physiological temperature as opposed to the common, lower experimental temperatures. At 37 degrees C, compared with 23 degrees C, the speed of both exocytosis and endocytosis was higher. The size of the recycling vesicle pool (in both number of vesicles and spatial extent) was twofold larger at 37 degrees C. In addition, although repeated 10 Hz electrical stimulation caused an NMDA receptor-dependent enlargement (averaging 170%) of the measurable recycling vesicle pool at 23 degrees C, the same stimulus repetition had no effect at 37 degrees C. These results show that it is potentially misleading to extend conclusions drawn about vesicle function or presynaptic plasticity at lowered experimental temperature to physiological conditions and that much new experimental work at the higher physiological temperature range will be needed to understand the true parameters of presynaptic functions.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources