

High-concentration rapid transients of glutamate mediate neural-glial communication via ectopic release
- PMID: 16107641
- PMCID: PMC6725396
- DOI: 10.1523/JNEUROSCI.1927-05.2005
High-concentration rapid transients of glutamate mediate neural-glial communication via ectopic release
Abstract
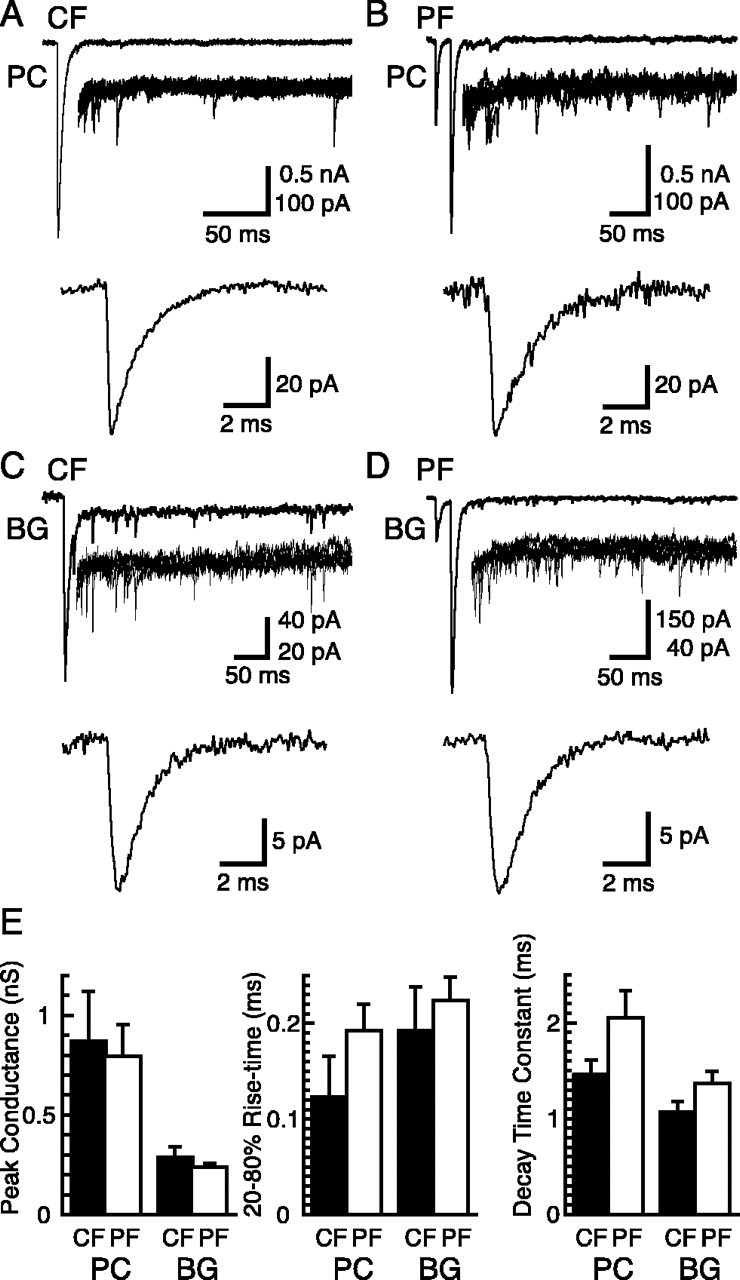
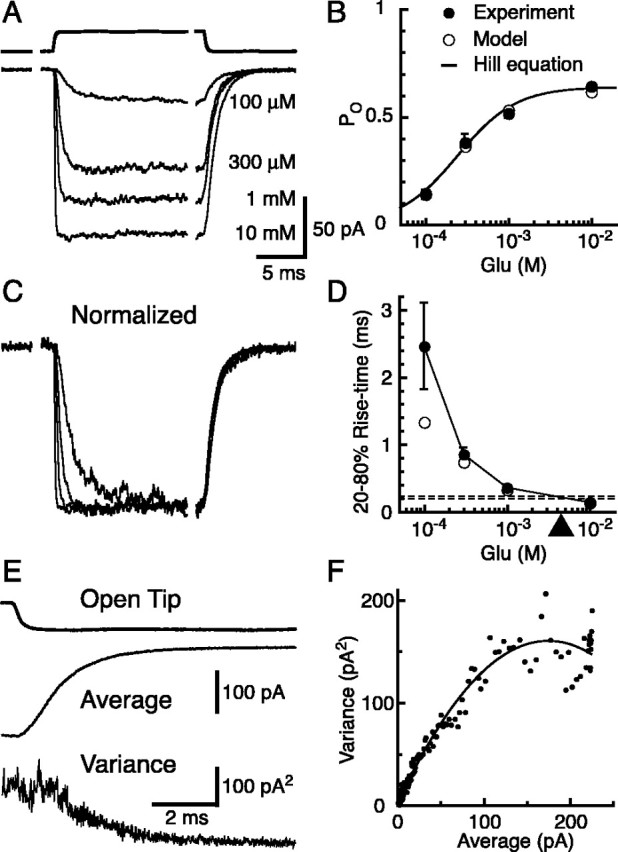
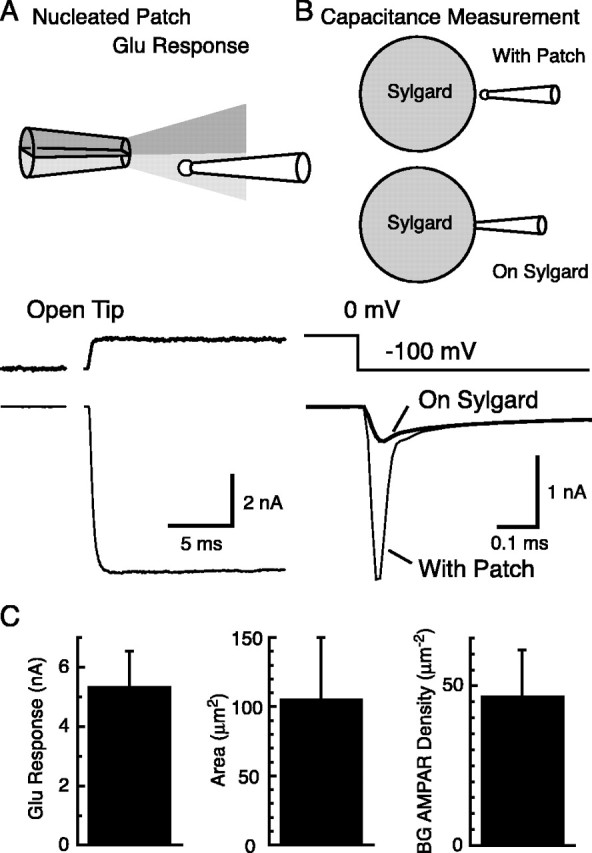
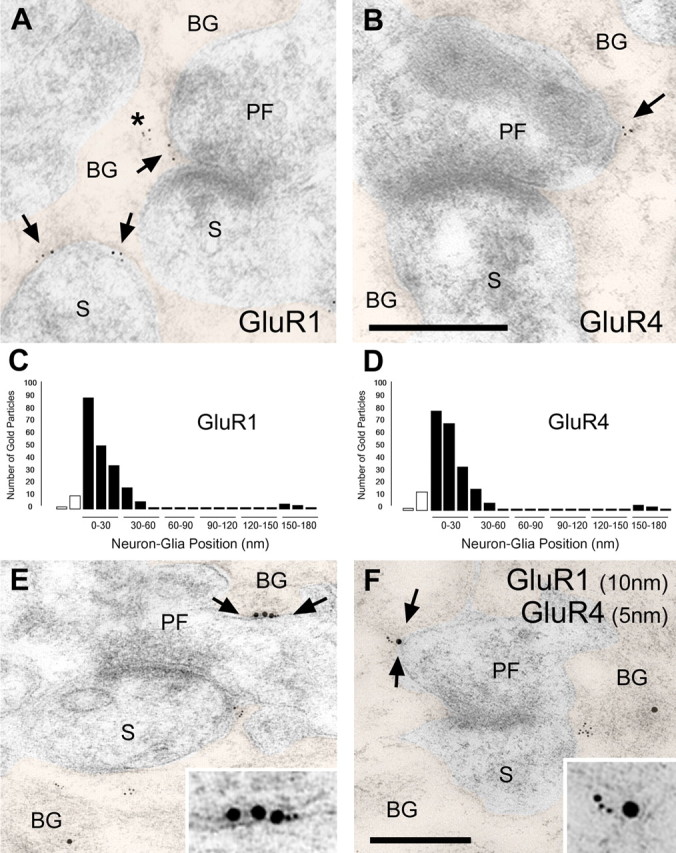
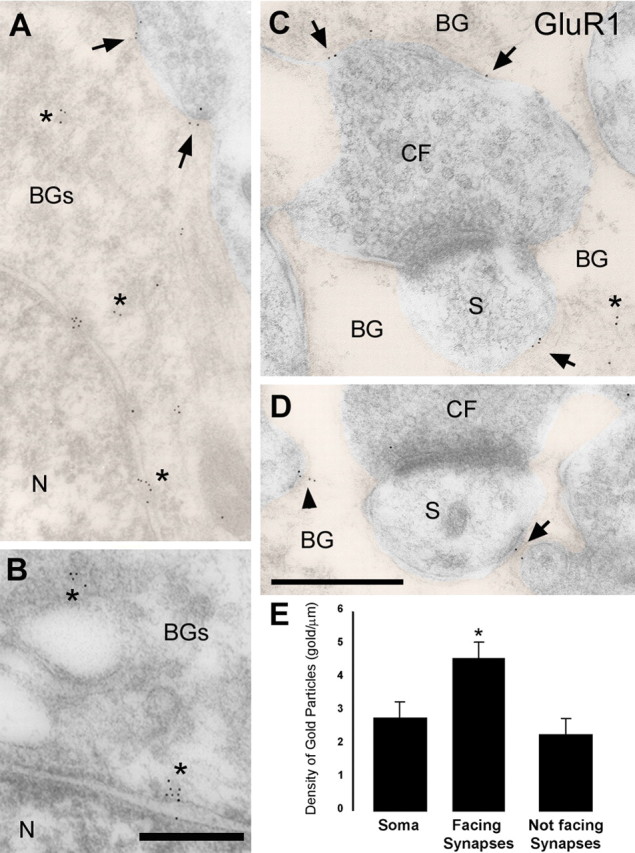
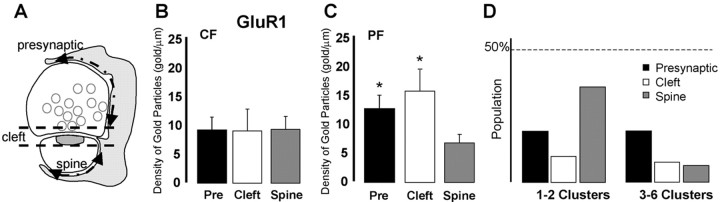
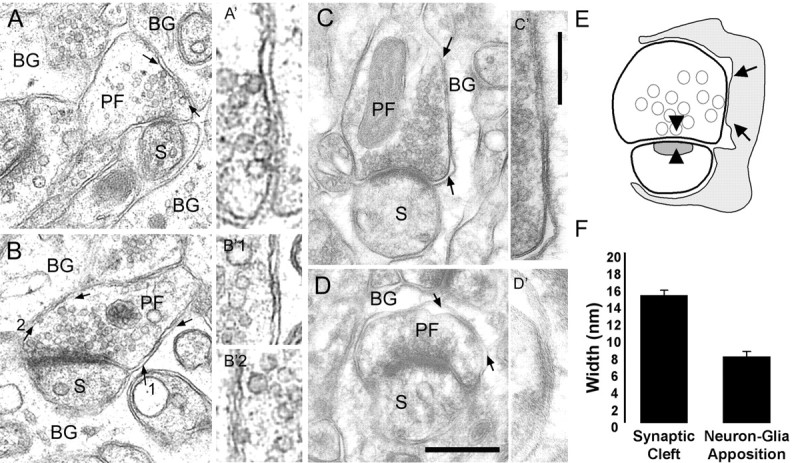
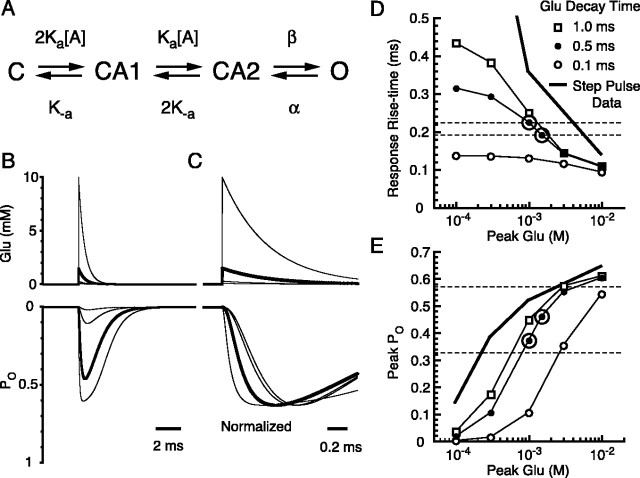
Until recently, communication from neurons to astrocytes was thought to be mediated by low-concentration transients of glutamate caused by spillover from the synaptic cleft. However, quantal events recorded in rat cerebellar Bergmann glial cells (BGs) have fast kinetics, comparable with those recorded in neurons. By combining outside-out patch recordings of BG AMPA receptors and quantitative electron microscopic analysis of glutamate receptor subunit 1 (GluR1) and GluR4 immunogold labeling measurements, at both the soma and membranes surrounding synapses, we estimate the absolute density of functional AMPA receptors. Using a kinetic model of BG AMPA receptors, we find that quantal events recorded in BGs are produced by high-concentration (approximately 1-1.5 mM), fast transients (approximately 0.5 ms decay) of glutamate, similar to transients within the synaptic cleft. Our results indicate that neural signaling to BGs is mediated by ectopic release of transmitter from presynaptic elements directly facing the BG membrane.
Figures
References
-
- Altman J, Bayer SA (1997) Development of cerebellar system in relation to its evolution, structure and functions. New York: CRC.
-
- Araque A, Carmignoto G, Haydon PG (2001) Dynamic signaling between astrocytes and neurons. Annu Rev Physiol 63: 795-813. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources