

Cationic sites on granzyme B contribute to cytotoxicity by promoting its uptake into target cells
- PMID: 16107729
- PMCID: PMC1190293
- DOI: 10.1128/MCB.25.17.7854-7867.2005
Cationic sites on granzyme B contribute to cytotoxicity by promoting its uptake into target cells
Abstract
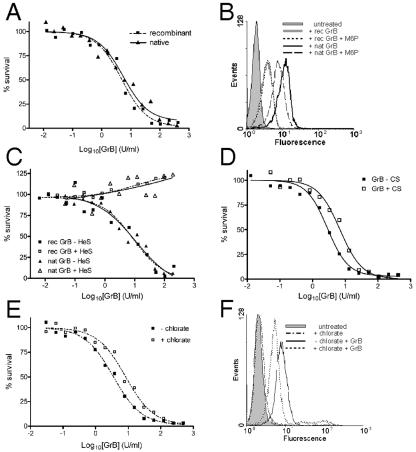
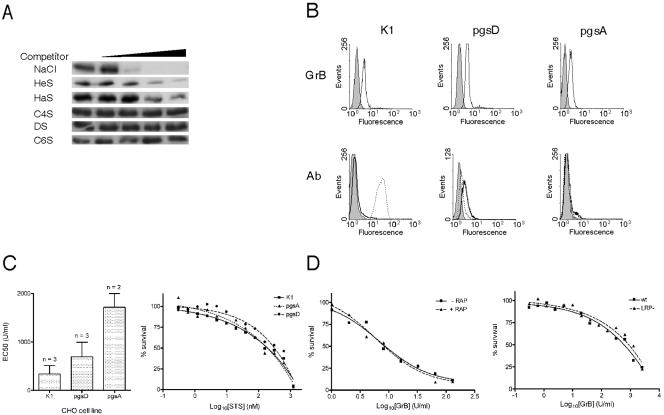
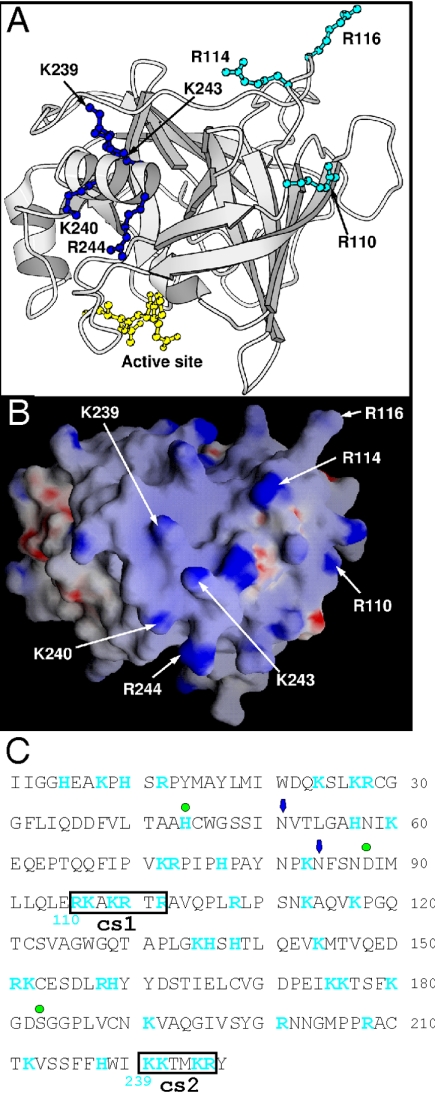
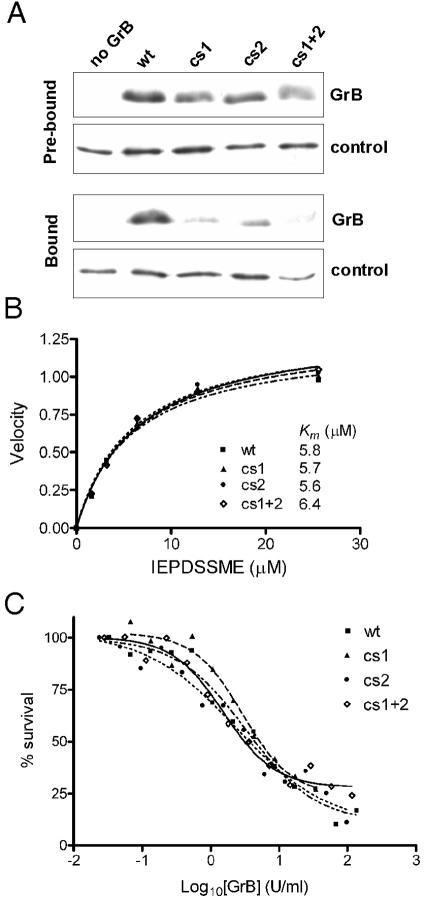
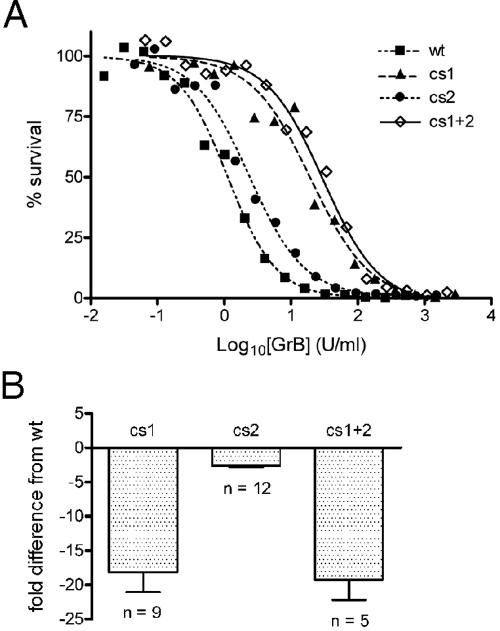
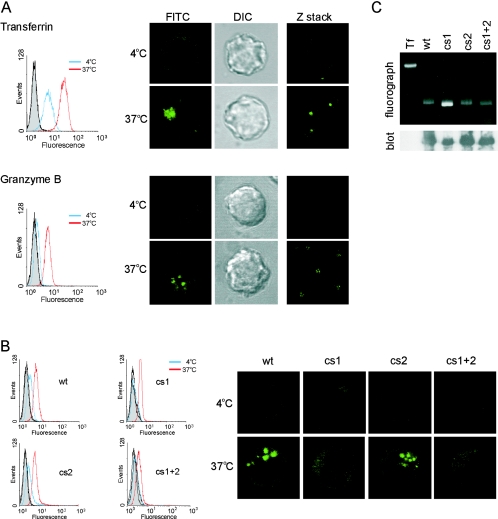
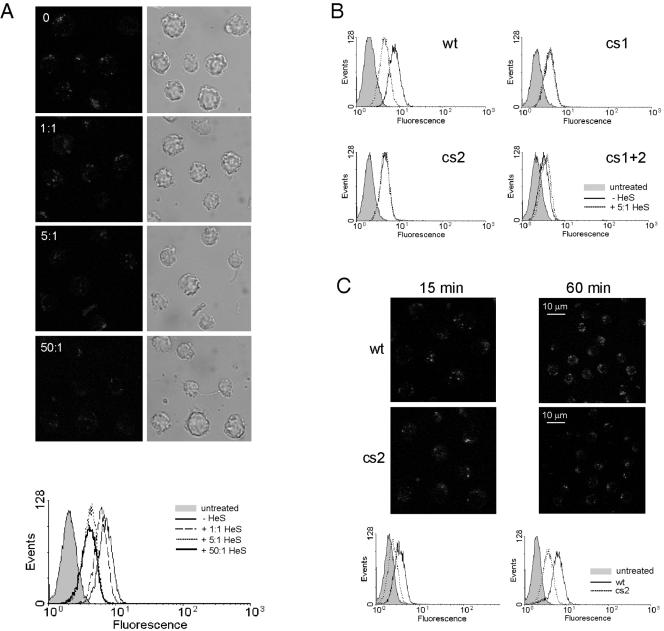
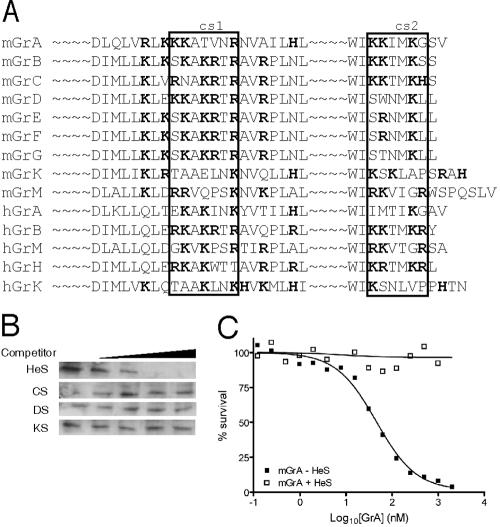
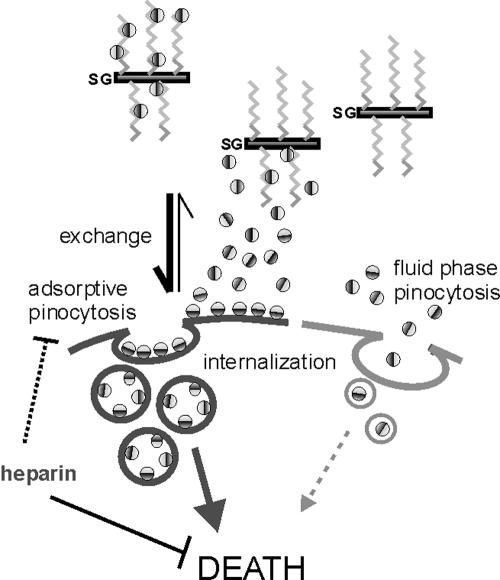
Granzyme B (GrB) is a key effector of cytotoxic lymphocyte-mediated cell death. It is delivered to target cells bound to the proteoglycan serglycin, but how it crosses the plasma membrane and accesses substrates in the cytoplasm is poorly understood. Here we identify two cationic sequences on GrB that facilitate its binding and uptake. Mutation of cationic sequence 1 (cs1) prevents accumulation of GrB in a distinctive intracellular compartment and reduces cytotoxicity 20-fold. Mutation of cs2 reduces accumulation in this intracellular compartment and cytotoxicity two- to threefold. We also show that GrB-mediated cytotoxicity is abrogated by heparin and that target cells deficient in cell surface sulfate or glycosaminoglycans resist GrB. However, heparin does not completely prevent GrB internalization and chondroitin 4-sulfate does not inhibit cytotoxicity, suggesting that glycosaminoglycans are not essential GrB receptors. We propose that GrB enters cells by nonselective adsorptive pinocytosis, exchanging from chondroitin sulfate on serglycin to anionic components of the cell surface. In this electrostatic "exchange-adsorption" model, cs1 and cs2 participate in binding of GrB to the cell surface, thereby promoting its uptake and eventual release into the cytoplasm.
Figures
References
-
- Baeuerle, P. A., and W. B. Huttner. 1986. Chlorate—a potent inhibitor of protein sulfation in intact cells. Biochem. Biophys. Res. Commun. 141:870-877. - PubMed
-
- Barry, M., and C. R. Bleackley. 2002. Cytotoxic lymphocytes: all roads lead to death. Nature Rev. Immunol. 2:401-409. - PubMed
-
- Bretthauer, R. K., and F. J. Castellino. 1997. Glycosylation of Pichia pastoris-derived proteins. Biotechnol. Appl. Biochem. 30:193-200. - PubMed
-
- Browne, K. A., E. Blink, V. R. Sutton, C. J. Froelich, D. A. Jans, and J. A. Trapani. 1999. Cytosolic delivery of granzyme B by bacterial toxins: evidence that endosomal disruption, in addition to transmembrane pore formation, is an important function of perforin. Mol. Cell. Biol. 19:8604-8615. - PMC - PubMed
-
- Cardin, A. D., and H. J. Weintraub. 1989. Molecular modeling of protein-glycosaminoglycan interactions. Arteriosclerosis 9:21-32. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous