

Mannheimia haemolytica leukotoxin induces apoptosis of bovine lymphoblastoid cells (BL-3) via a caspase-9-dependent mitochondrial pathway
- PMID: 16113266
- PMCID: PMC1231077
- DOI: 10.1128/IAI.73.9.5504-5513.2005
Mannheimia haemolytica leukotoxin induces apoptosis of bovine lymphoblastoid cells (BL-3) via a caspase-9-dependent mitochondrial pathway
Abstract
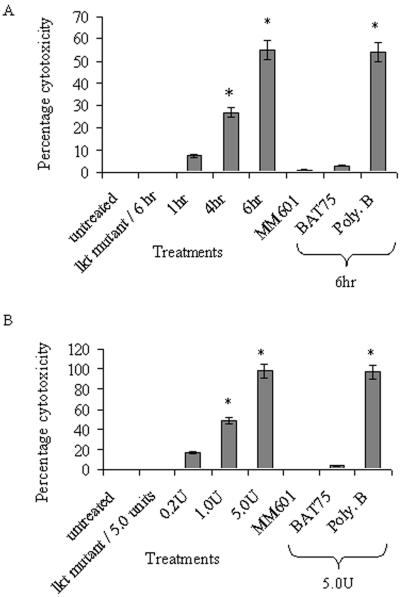
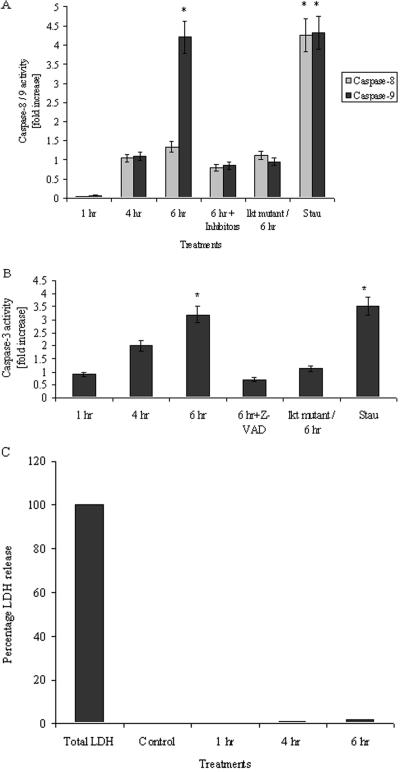
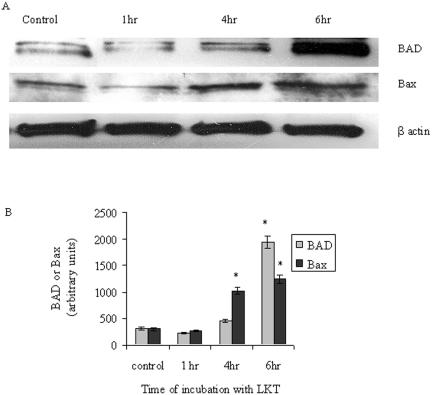
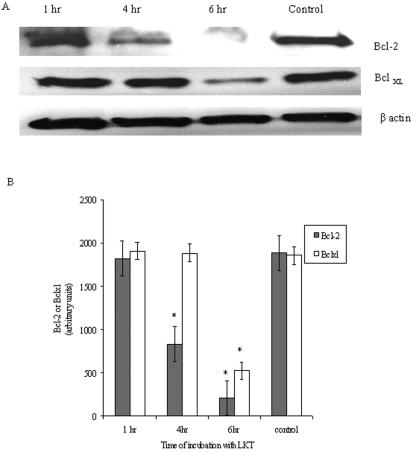
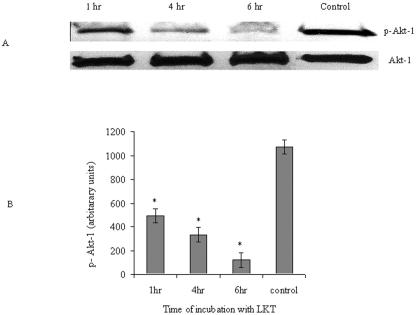
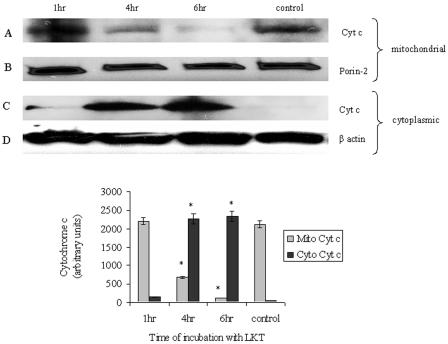
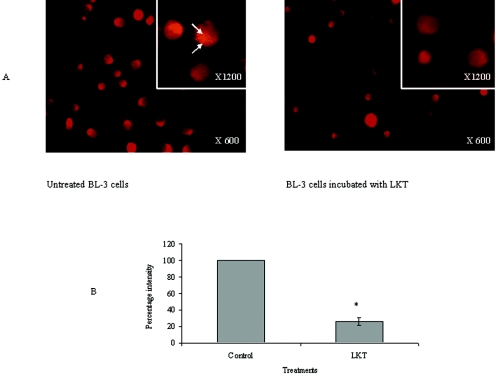
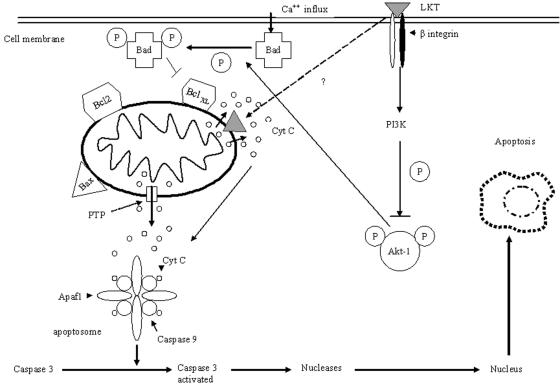
Mannheimia haemolytica is a key pathogen in the bovine respiratory disease complex. It produces a leukotoxin (LKT) that is an important virulence factor, causing cell death in bovine leukocytes. The LKT binds to the beta(2) integrin CD11a/CD18, which usually activates signaling pathways that facilitate cell survival. In this study, we investigated mechanisms by which LKT induces death in bovine lymphoblastoid cells (BL-3). Incubation of BL-3 cells with a low concentration of LKT results in the activation of caspase-3 and caspase-9 but not caspase-8. Similarly, the proapoptotic proteins Bax and BAD were significantly elevated, while the antiapoptotic proteins Bcl-2, Bcl(XL) and Akt-1 were downregulated. Following exposure to LKT, we also observed a reduction in mitochondrial cytochrome c and corresponding elevation of cytosolic cytochrome c, suggesting translocation from the mitochondrial compartment to the cytosol. Consistent with this observation, tetramethylrhodamine ethyl ester perchlorate staining revealed that mitochondrial membrane potential was significantly reduced. These data suggest that LKT induces apoptosis of BL-3 cells via a caspase-9-dependent mitochondrial pathway. Furthermore, scanning electron micrographs of mitochondria from LKT-treated BL-3 cells revealed lesions in the outer mitochondrial membrane, which are larger than previous reports of the permeability transition pore through which cytochrome c is usually released.
Figures

References
-
- Ambagala, T. C., A. P. Ambagala, and S. Srikumaran. 1999. The leukotoxin of Pasteurella haemolytica binds to beta(2) integrins on bovine leukocytes. FEMS Microbiol Lett. 179:161-167. - PubMed
-
- Antonsson, B. 2004. Mitochondria and the Bcl-2 family proteins in apoptosis signaling pathways. Mol. Cell. Biochem. 256-257:141-155. - PubMed
-
- Araya, R., T. Uehara, and Y. Nomura. 1998. Hypoxia induces apoptosis in human neuroblastoma SK-N-MC cells by caspase activation accompanying cytochrome c release from mitochondria. FEBS Lett. 439:168-172. - PubMed
-
- Aronis, A., J. A. Melendez, O. Golan, S. Shilo, N. Dicter, and O. Tirosh. 2003. Potentiation of Fas-mediated apoptosis by attenuated production of mitochondria-derived reactive oxygen species. Cell Death Differ. 10:335-344. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
