

Tracking antigen-specific CD8 T lymphocytes in the lungs of mice vaccinated with the Mtb72F polyprotein
- PMID: 16113299
- PMCID: PMC1231129
- DOI: 10.1128/IAI.73.9.5809-5816.2005
Tracking antigen-specific CD8 T lymphocytes in the lungs of mice vaccinated with the Mtb72F polyprotein
Abstract
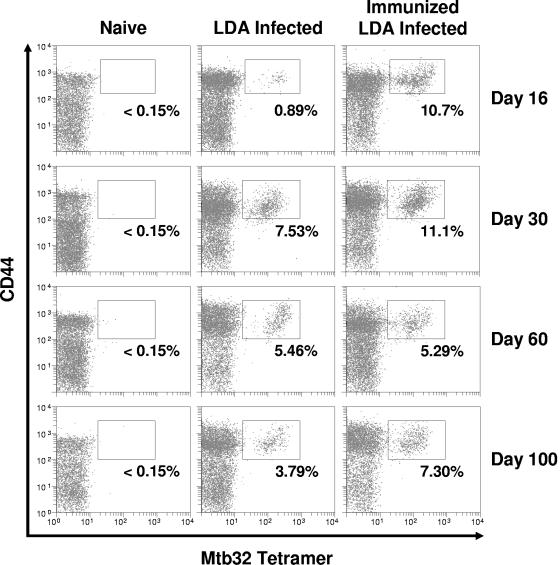
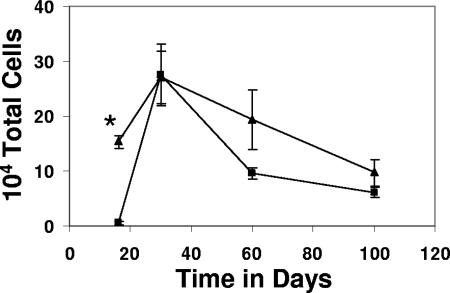
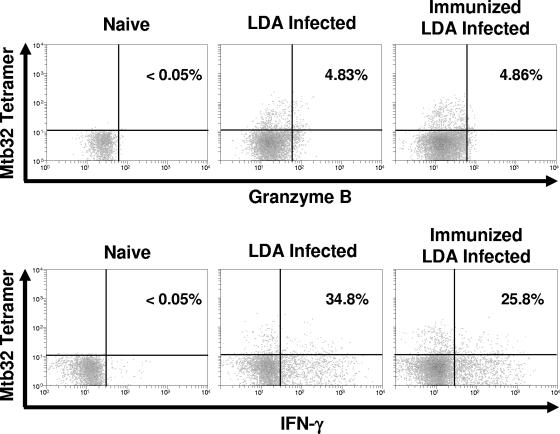
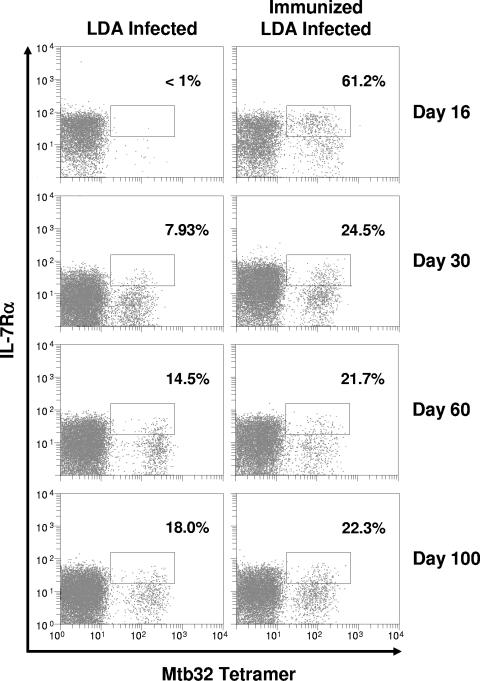
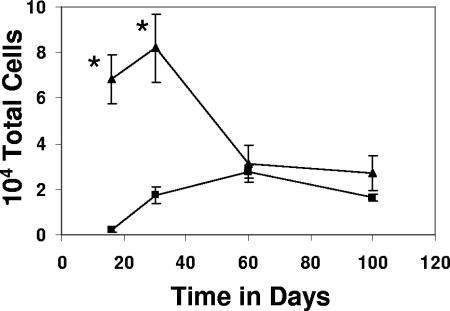
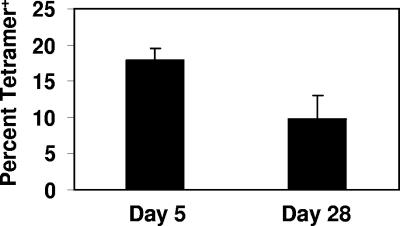
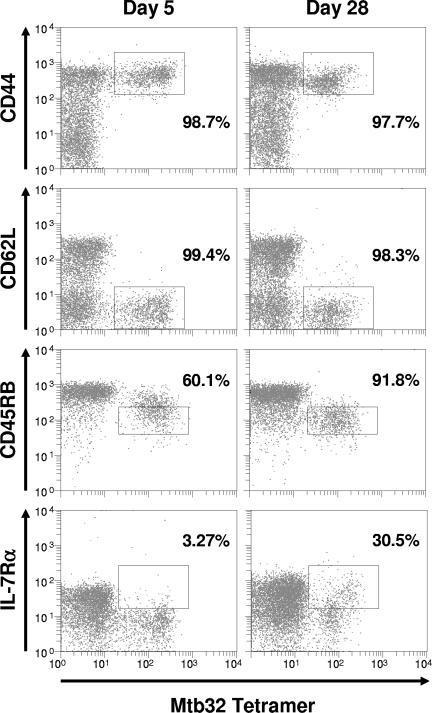
This study used a major histocompatibility complex class I tetramer reagent to track antigen-specific CD8 T cells in the lungs of mice immunized with the tuberculosis vaccine candidate Mtb72F. The results show that CD8 T cells recognizing an immunodominant Mtb32-specific epitope could be detected in significant numbers over the course of infection in mice exposed to low-dose aerosol challenge with Mycobacterium tuberculosis and that prior vaccination substantially increased the numbers of these cells early in the lungs. The effector phenotype of the cells was shown by the demonstration that many secreted gamma interferon, but very few contained granzyme B. As the course of the infection progressed, many activated CD8 T cells down-regulated expression of CD45RB and upregulated expression of the interleukin-7 receptor alpha chain, indicating a transition of these cells to a state of memory. These data support the hypothesis that M. tuberculosis-specific CD8 T cells can be targeted by vaccination with the Mtb72F polyprotein.
Figures
References
-
- Andersen, P. 2001. TB vaccines: progress and problems. Trends Immunol. 22:160-168. - PubMed
-
- Bloom, B. R., and C. J. Murray. 1992. Tuberculosis: commentary on a reemergent killer. Science 257:1055-1064. - PubMed
-
- Boom, W. H. 1996. The role of T-cell subsets in Mycobacterium tuberculosis infection. Infect. Agents Dis. 5:73-81. - PubMed
-
- Brandt, L., Y. A. Skeiky, M. R. Alderson, Y. Lobet, W. Dalemans, O. C. Turner, R. J. Basaraba, A. A. Izzo, T. M. Lasco, P. L. Chapman, S. G. Reed, and I. M. Orme. 2004. The protective effect of the Mycobacterium bovis BCG vaccine is increased by coadministration with the Mycobacterium tuberculosis 72-kilodalton fusion polyprotein Mtb72F in M. tuberculosis-infected guinea pigs. Infect. Immun. 72:6622-6632. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
