

NKT cell stimulation with glycolipid antigen in vivo: costimulation-dependent expansion, Bim-dependent contraction, and hyporesponsiveness to further antigenic challenge
- PMID: 16116198
- PMCID: PMC1360163
- DOI: 10.4049/jimmunol.175.5.3092
NKT cell stimulation with glycolipid antigen in vivo: costimulation-dependent expansion, Bim-dependent contraction, and hyporesponsiveness to further antigenic challenge
Abstract
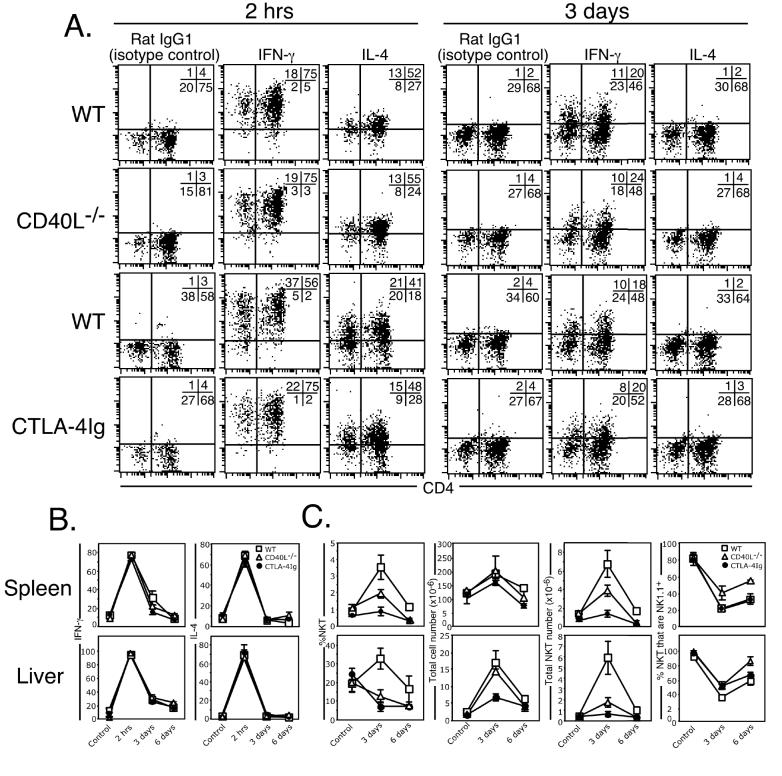
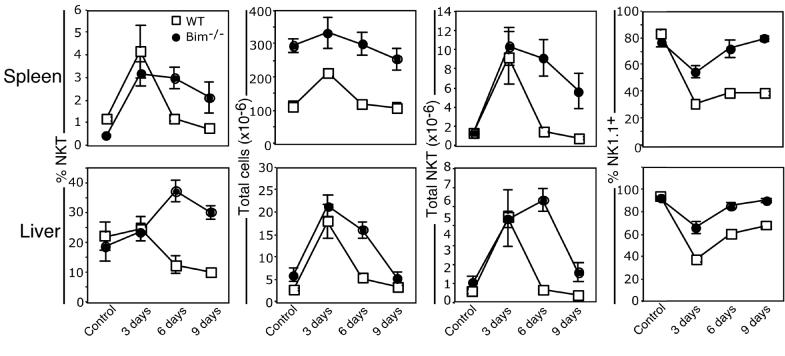
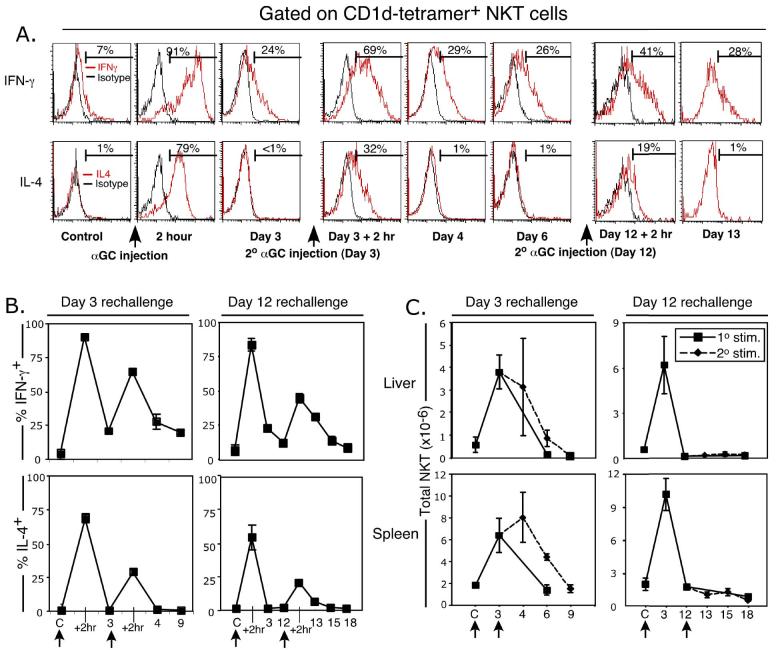
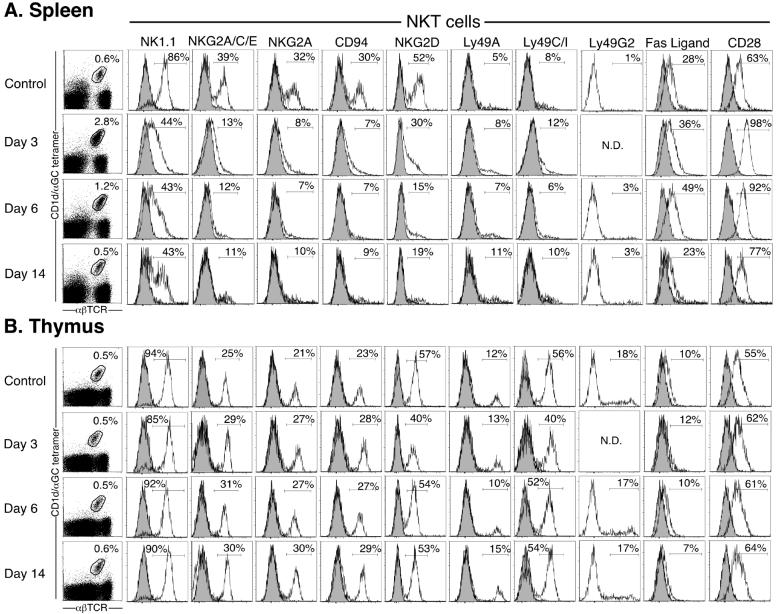
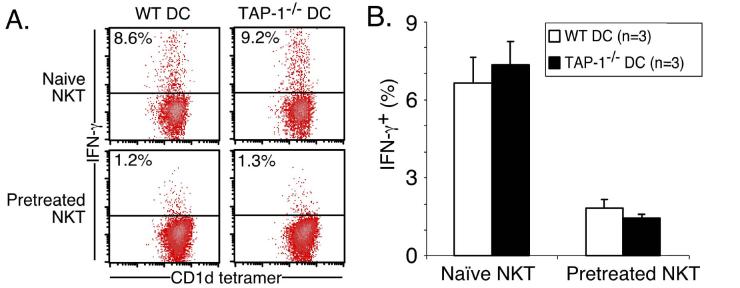
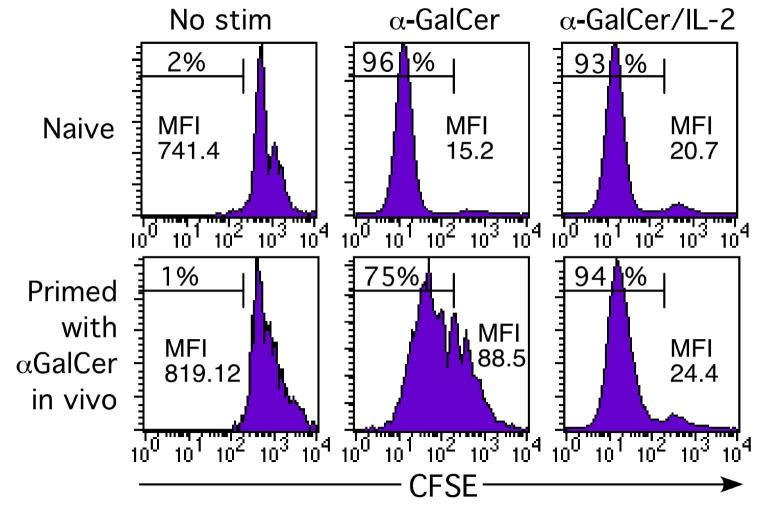
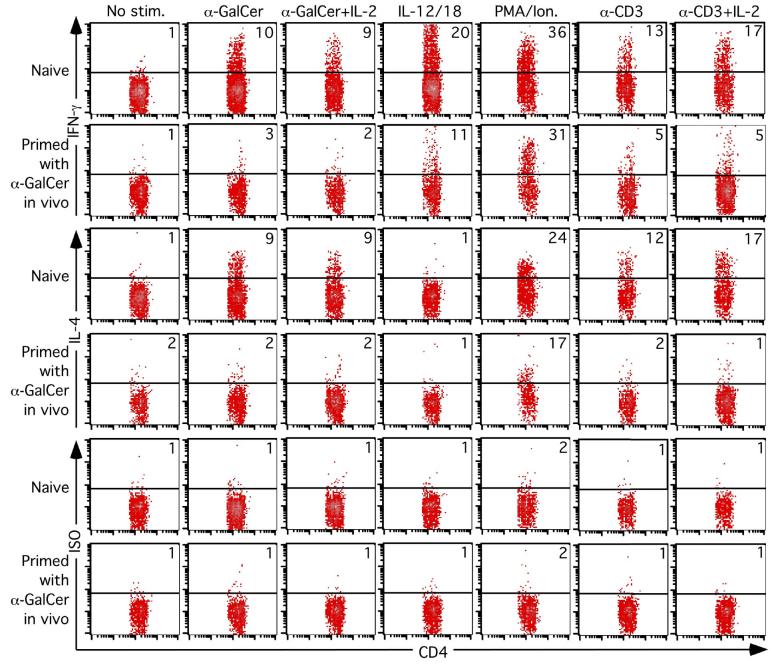
Activation of NKT cells using the glycolipid alpha-galactosylceramide (alpha-GalCer) has availed many investigations into their immunoregulatory and therapeutic potential. However, it remains unclear how they respond to stimulation in vivo, which costimulatory pathways are important, and what factors (e.g., Ag availability and activation-induced cell death) limit their response. We have explored these questions in the context of an in vivo model of NKT cell dynamics spanning activation, population expansion, and subsequent contraction. Neither the B7/CD28 nor the CD40/CD40L costimulatory pathway was necessary for cytokine production by activated NKT cells, either early (2 h) or late (3 days) after initial stimulation, but both pathways were necessary for normal proliferative expansion of NKT cells in vivo. The proapoptotic Bcl-2 family member Bim was necessary for normal contraction of the NKT cell population between days 3-9 after stimulation, suggesting that the pool size is regulated by apoptotic death, similar to that of conventional T cells. Ag availability was not the limiting factor for NKT cell expansion in vivo, and a second alpha-GalCer injection induced a very blunted response, whereby cytokine production was reduced and further expansion did not occur. This appeared to be a form of anergy that was intrinsic to NKT cells and was not associated with inhibitory NK receptor signaling. Furthermore, NKT cells from mice pre-challenged with alpha-GalCer in vivo showed little cytokine production and reduced proliferation in vitro. In summary, this study significantly enhances our understanding of how NKT cells respond to primary and secondary antigenic challenge in vivo.
Figures
References
-
- Godfrey DI, MacDonald HR, Kronenberg M, Smyth MJ, Van Kaer L. NKT cells: what's in a name? Nat Rev Immunol. 2004;4:231. - PubMed
-
- Brigl M, Brenner MB. CD1: Antigen Presentation and T Cell Function. Annu Rev Immunol. 2004;22:817. - PubMed
-
- Kronenberg M. Toward an understanding of NKT cell biology: Progress and Paradoxes. Annu Rev Immunol. 2005;23:877. - PubMed
-
- Hayakawa Y, Godfrey DI, Smyth MJ. Alpha-galactosylceramide: potential immunomodulatory activity and future application. Curr Med Chem. 2004;11:241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
