

A human ortholog of archaeal DNA repair protein Hef is defective in Fanconi anemia complementation group M
- PMID: 16116422
- PMCID: PMC2704909
- DOI: 10.1038/ng1626
A human ortholog of archaeal DNA repair protein Hef is defective in Fanconi anemia complementation group M
Abstract
Fanconi anemia is a genetic disease characterized by genomic instability and cancer predisposition. Nine genes involved in Fanconi anemia have been identified; their products participate in a DNA damage-response network involving BRCA1 and BRCA2 (refs. 2,3). We previously purified a Fanconi anemia core complex containing the FANCL ubiquitin ligase and six other Fanconi anemia-associated proteins. Each protein in this complex is essential for monoubiquitination of FANCD2, a key reaction in the Fanconi anemia DNA damage-response pathway. Here we show that another component of this complex, FAAP250, is mutant in individuals with Fanconi anemia of a new complementation group (FA-M). FAAP250 or FANCM has sequence similarity to known DNA-repair proteins, including archaeal Hef, yeast MPH1 and human ERCC4 or XPF. FANCM can dissociate DNA triplex, possibly owing to its ability to translocate on duplex DNA. FANCM is essential for monoubiquitination of FANCD2 and becomes hyperphosphorylated in response to DNA damage. Our data suggest an evolutionary link between Fanconi anemia-associated proteins and DNA repair; FANCM may act as an engine that translocates the Fanconi anemia core complex along DNA.
Conflict of interest statement
Figures
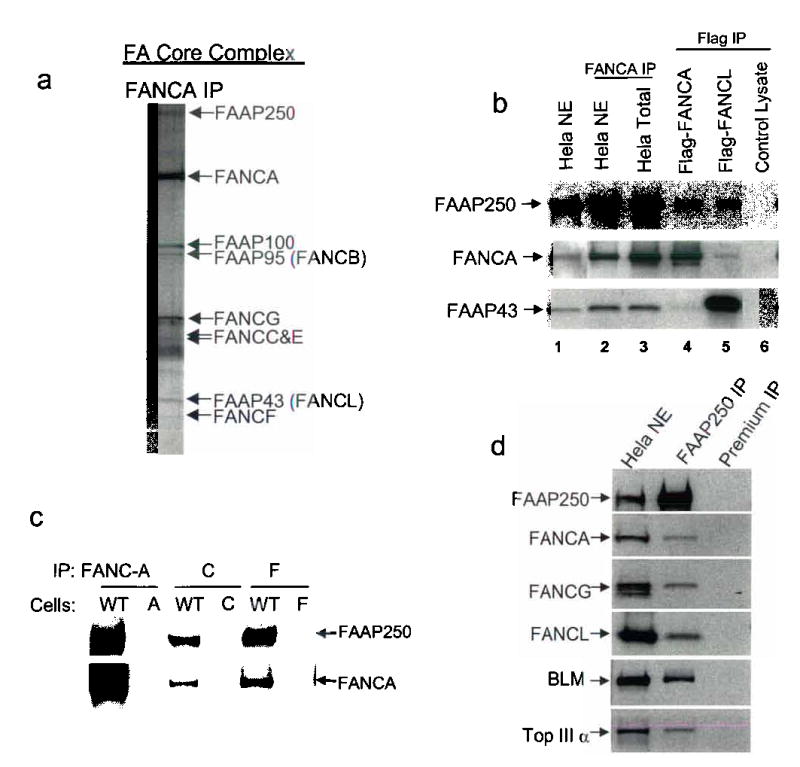
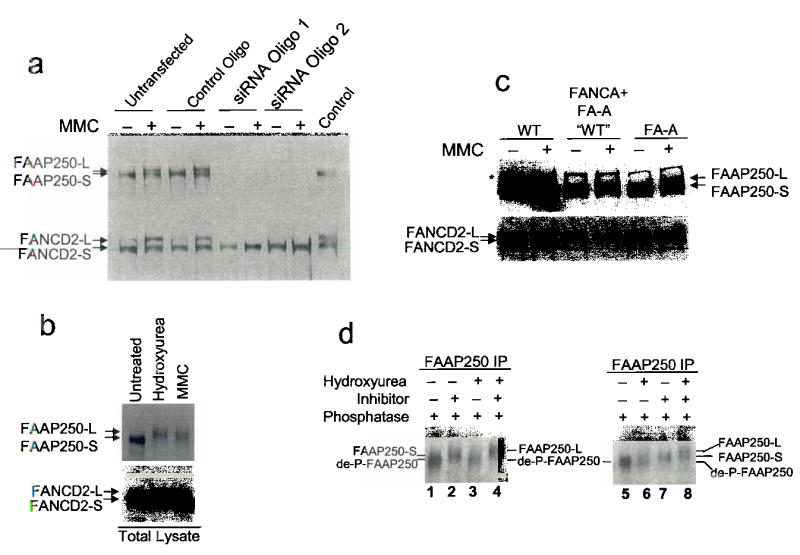
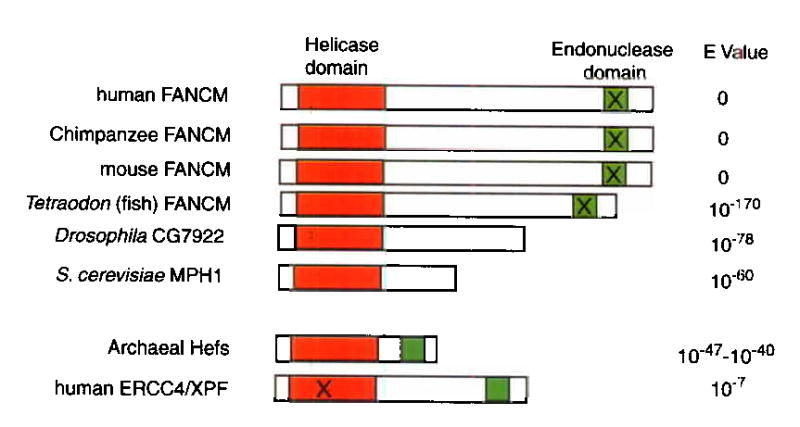
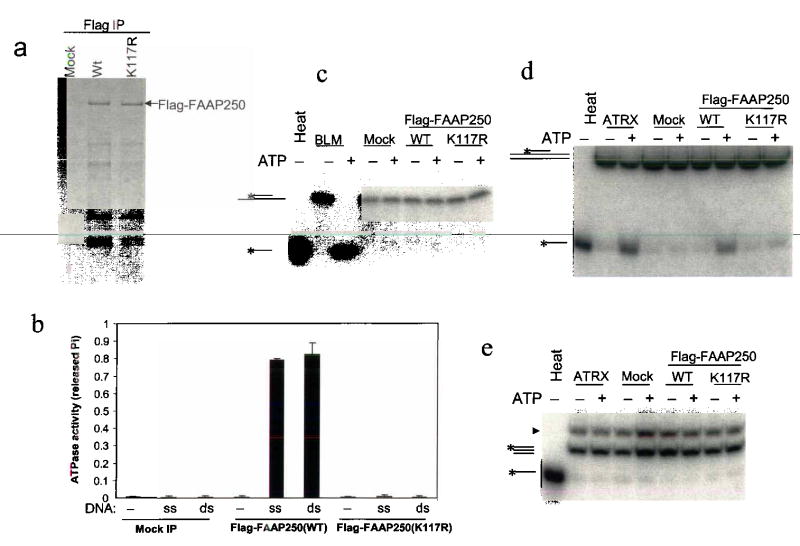
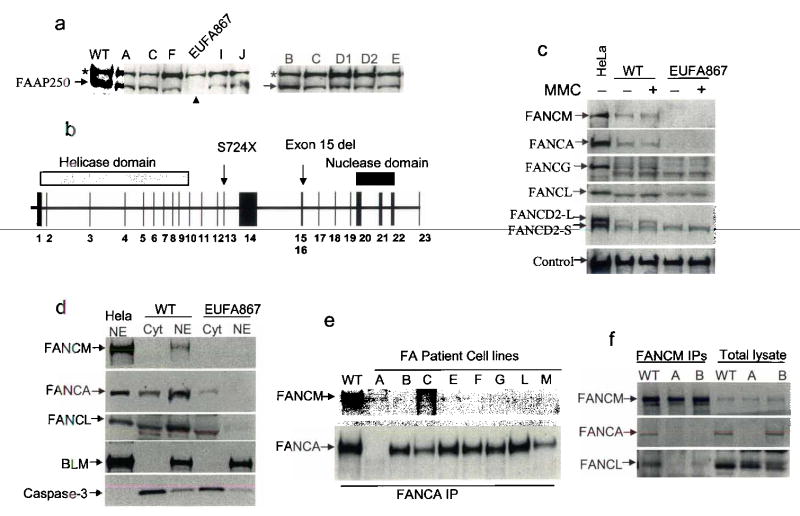
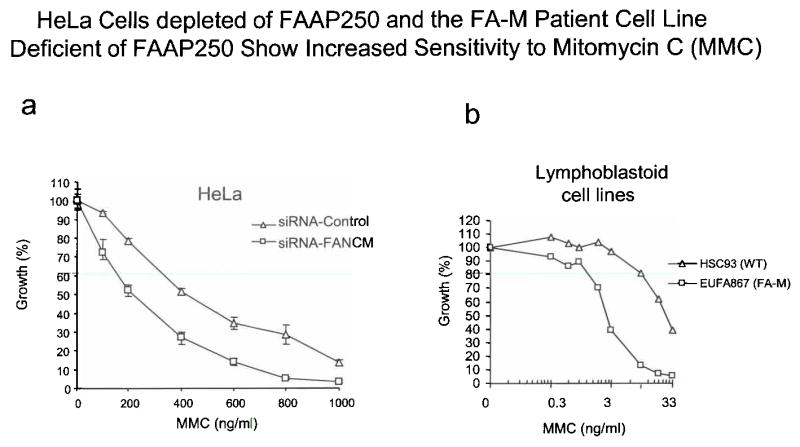


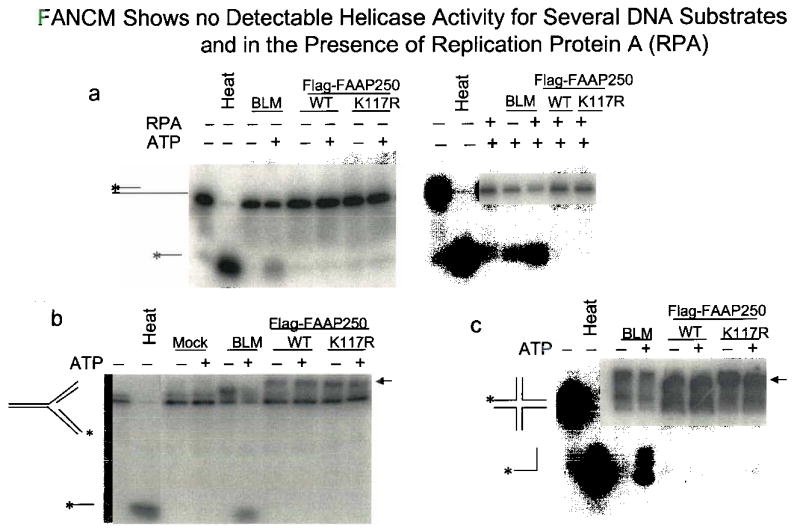
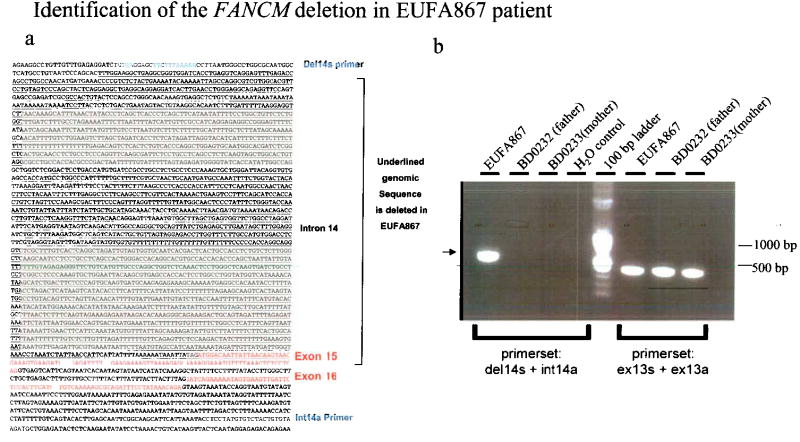
Comment in
-
Unraveling the Fanconi anemia-DNA repair connection.Nat Genet. 2005 Sep;37(9):921-2. doi: 10.1038/ng0905-921. Nat Genet. 2005. PMID: 16132046 No abstract available.
References
-
- Joenje H, Patel KJ. The emerging genetic and molecular basis of fanconi anaemia. Nat Rev Genet. 2001;2:446–59. - PubMed
-
- Garcia-Higuera I, et al. Interaction of the Fanconi anemia proteins and BRCA1 in a common pathway. Mol Cell. 2001;7:249–62. - PubMed
-
- Howlett NG, et al. Biallelic Inactivation of BRCA2 in Fanconi Anemia. Science. 2002;13:13. - PubMed
-
- Meetei AR, et al. A novel ubiquitin ligase is deficient in Fanconi anemia. Nat Genet. 2003;35:165–70. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
