

An astrocytic basis of epilepsy
- PMID: 16116433
- PMCID: PMC1850946
- DOI: 10.1038/nm1277
An astrocytic basis of epilepsy
Abstract
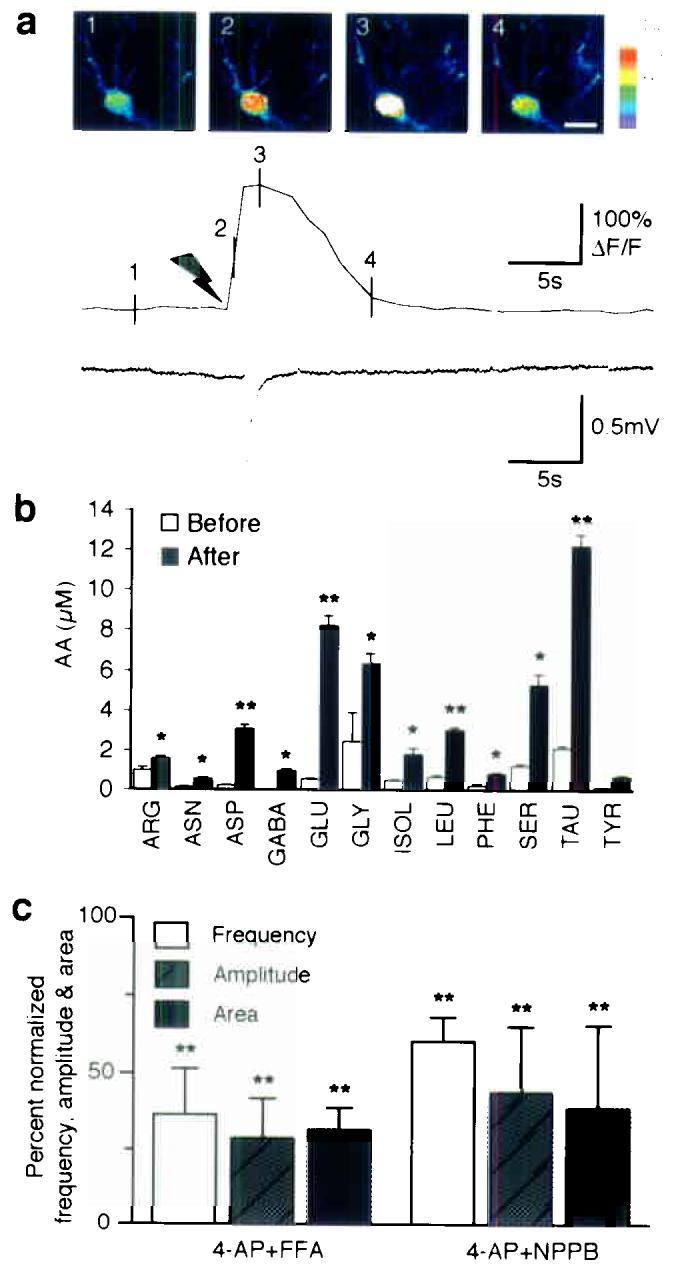
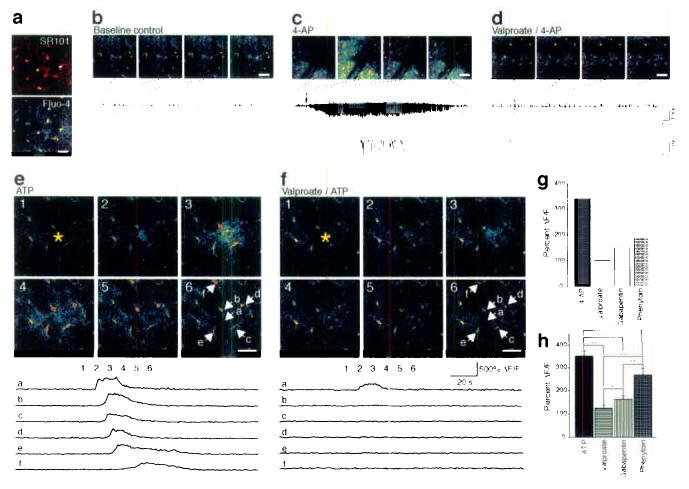
Hypersynchronous neuronal firing is a hallmark of epilepsy, but the mechanisms underlying simultaneous activation of multiple neurons remains unknown. Epileptic discharges are in part initiated by a local depolarization shift that drives groups of neurons into synchronous bursting. In an attempt to define the cellular basis for hypersynchronous bursting activity, we studied the occurrence of paroxysmal depolarization shifts after suppressing synaptic activity using tetrodotoxin (TTX) and voltage-gated Ca(2+) channel blockers. Here we report that paroxysmal depolarization shifts can be initiated by release of glutamate from extrasynaptic sources or by photolysis of caged Ca(2+) in astrocytes. Two-photon imaging of live exposed cortex showed that several antiepileptic agents, including valproate, gabapentin and phenytoin, reduced the ability of astrocytes to transmit Ca(2+) signaling. Our results show an unanticipated key role for astrocytes in seizure activity. As such, these findings identify astrocytes as a proximal target for the treatment of epileptic disorders.
Figures
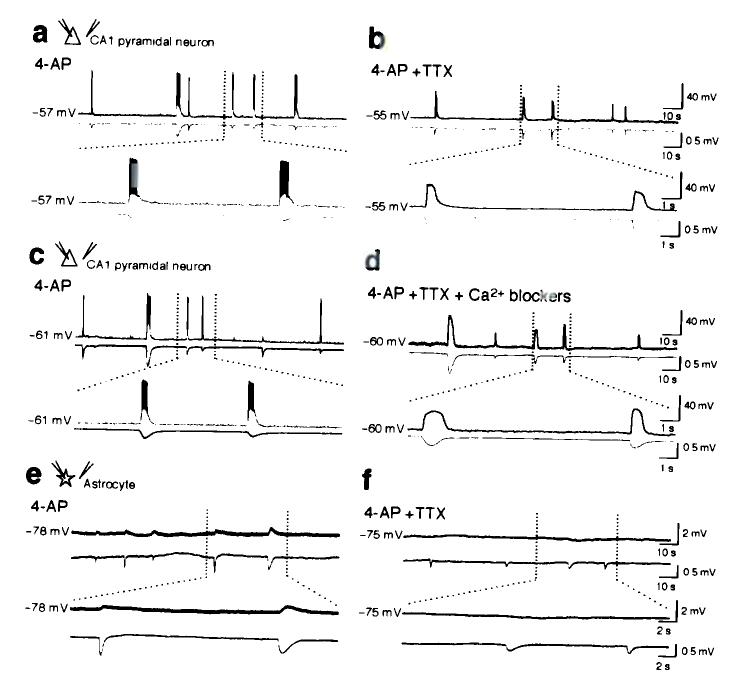
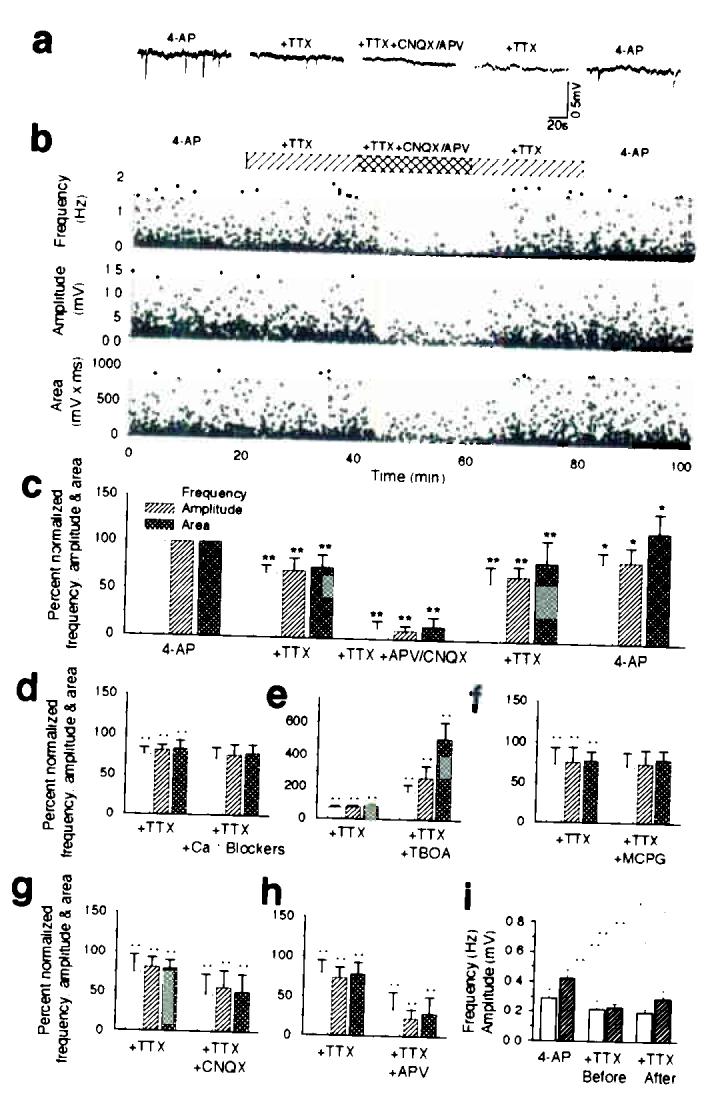
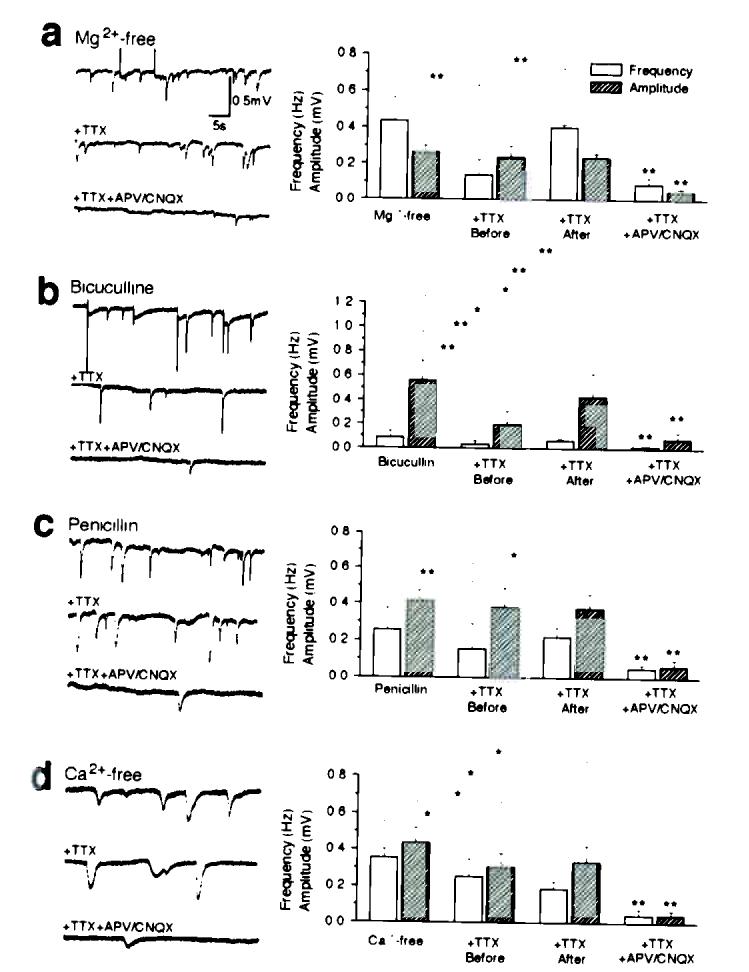
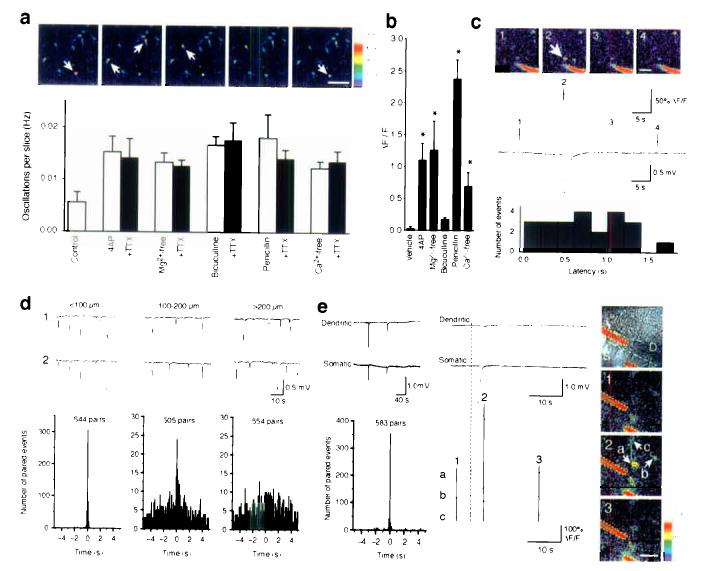
Comment in
-
Astrocytes get in the act in epilepsy.Nat Med. 2005 Sep;11(9):919-20. doi: 10.1038/nm0905-919. Nat Med. 2005. PMID: 16145568 Free PMC article.
-
Does glutamate released by astrocytes cause focal epilepsy?Epilepsy Curr. 2006 Sep-Oct;6(5):173-6. doi: 10.1111/j.1535-7511.2006.00136.x. Epilepsy Curr. 2006. PMID: 17260050 Free PMC article. No abstract available.
References
-
- Wyllie E. The treatment of epilepsy Principles and Practice. lippincot, Williams, and Wilkins; New York: 2001.
-
- Heinemann U, Gabriel S, Schuchmann S, Eder C. Contribution of astrocytes to seizure activity. Adv. Neurol. 1999;79:583–590. - PubMed
-
- Rogawski MA, Loscher W. The neurobiology of antiepileptic drugs. Nat. Rev. Neurosci. 2004;5:553–564. - PubMed
-
- Meldrum BS. Update on the mechanism of action of antiepileptic drugs. Epilepsia. 1996;37(Suppl. 6):S4–11. - PubMed
-
- Parpura V, et al. Glutamate-mediated astrocyte-neuron signalling. Nature. 1994;369:744–747. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
