

Bone marrow dysfunction in mice lacking the cytokine receptor gp130 in endothelial cells
- PMID: 16118327
- PMCID: PMC1895244
- DOI: 10.1182/blood-2005-02-0671
Bone marrow dysfunction in mice lacking the cytokine receptor gp130 in endothelial cells
Abstract
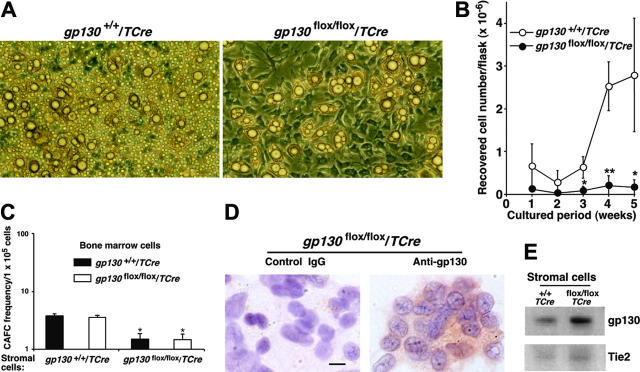
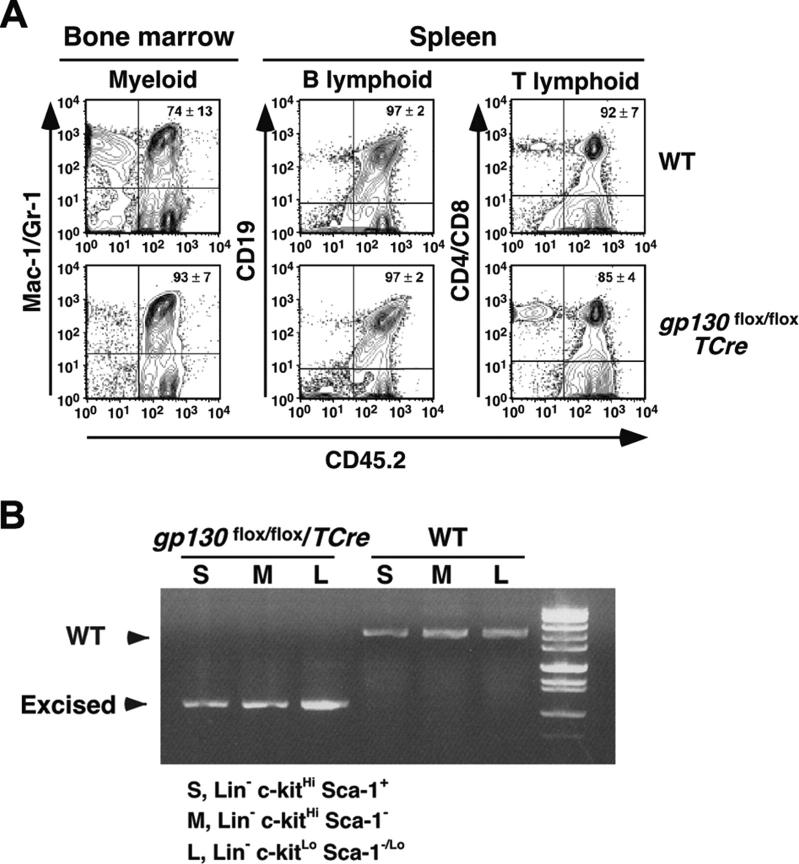
In vitro studies suggest that bone marrow endothelial cells contribute to multilineage hematopoiesis, but this function has not been studied in vivo. We used a Cre/loxP-mediated recombination to produce mice that lacked the cytokine receptor subunit gp130 in hematopoietic and endothelial cells. Although normal at birth, the mice developed bone marrow dysfunction that was accompanied by splenomegaly caused by extramedullary hematopoiesis. The hypocellular marrow contained myeloerythroid progenitors and functional repopulating stem cells. However, long-term bone marrow cultures produced few hematopoietic cells despite continued expression of gp130 in most stromal cells. Transplanting gp130-deficient bone marrow into irradiated wild-type mice conferred normal hematopoiesis, whereas transplanting wild-type bone marrow into irradiated gp130-deficient mice did not cure the hematopoietic defects. These data provide evidence that gp130 expression in the bone marrow microenvironment, most likely in endothelial cells, makes an important contribution to hematopoiesis.
Figures
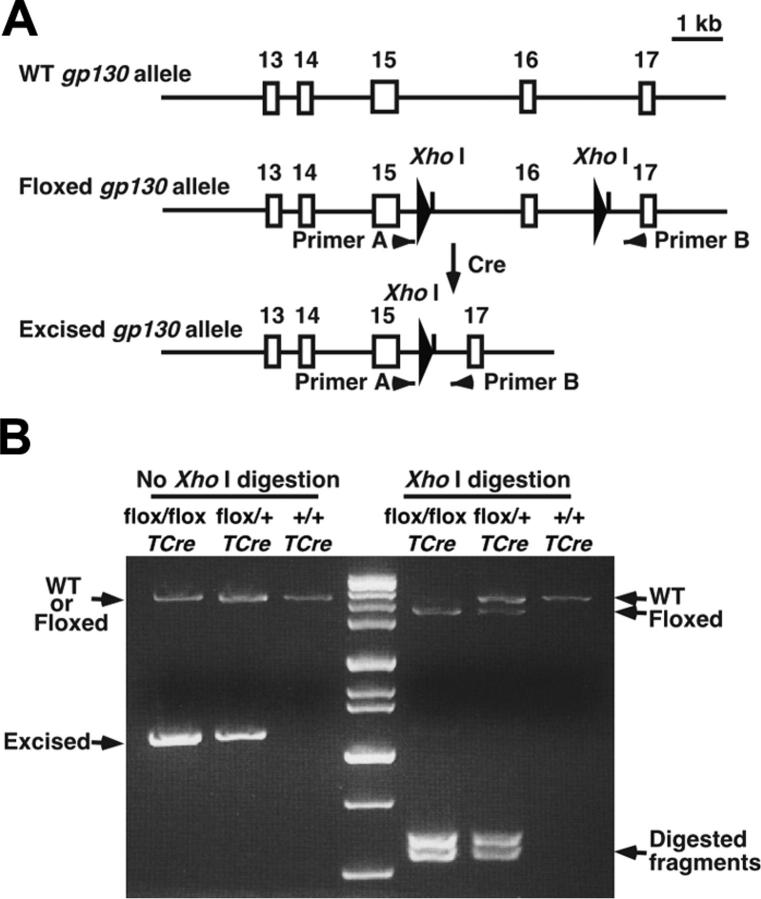
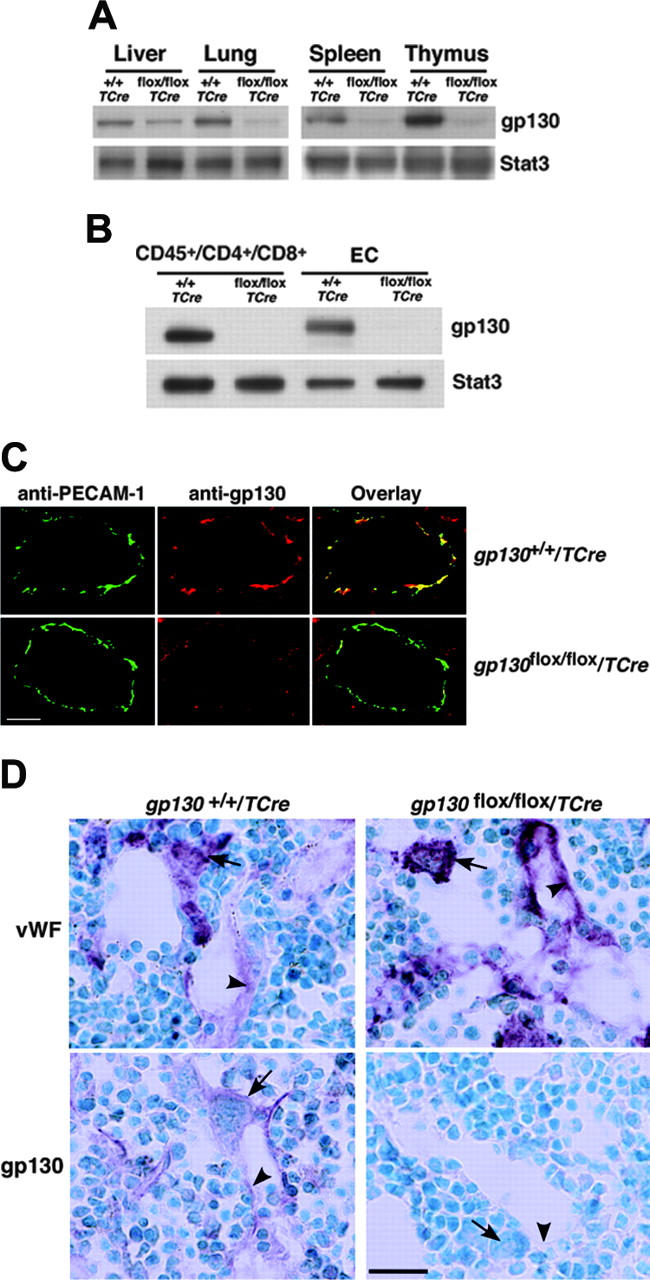
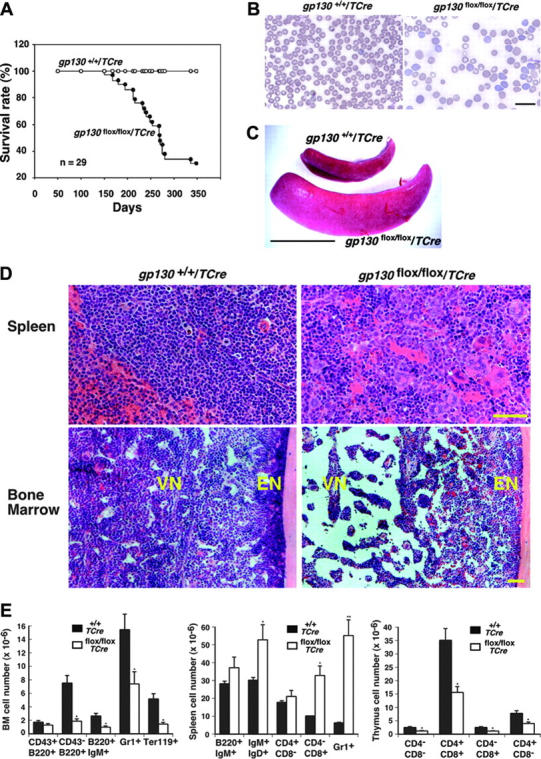
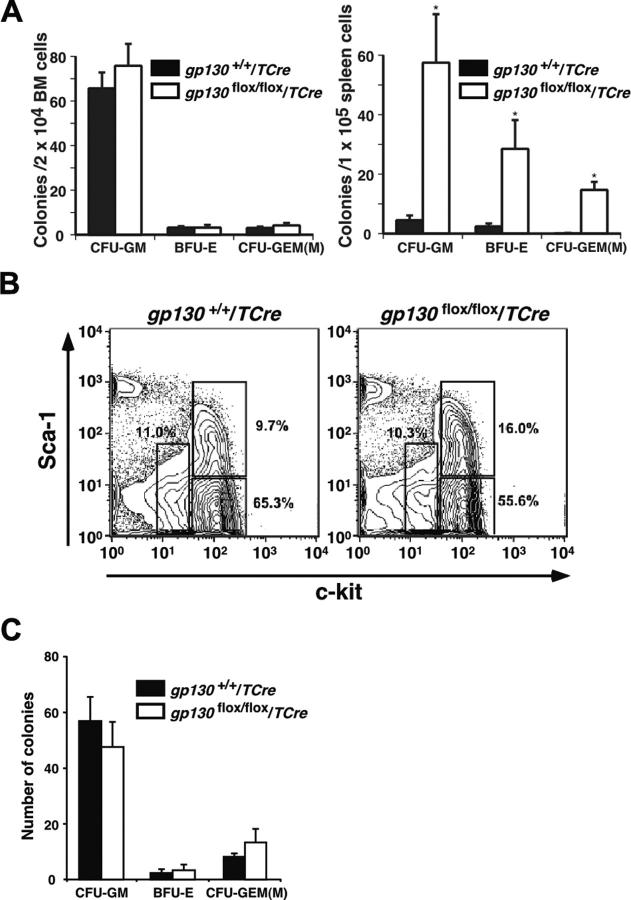
References
-
- Kamimura D, Ishihara K, Hirano T. IL-6 signal transduction and its physiological roles: the signal orchestration model. Rev Physiol Biochem Pharmacol. 2003;149: 1-38. - PubMed
-
- Taga T, Kishimoto T. Gp130 and the interleukin-6 family of cytokines. Annu Rev Immunol. 1997;15: 797-819. - PubMed
-
- Hirota H, Chen J, Betz UAK, et al. Loss of a gp130 cardiac muscle cell survival pathway is a critical event in the onset of heart failure during biomechanical stress. Cell. 1999;97: 189-198. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
