

The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis
- PMID: 16126837
- PMCID: PMC1242265
- DOI: 10.1105/tpc.105.033415
The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis
Abstract
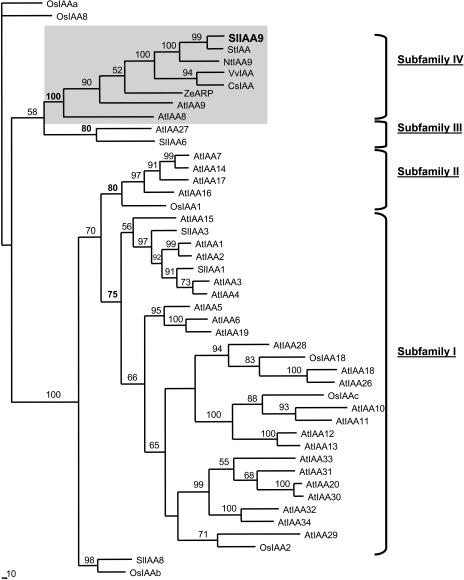
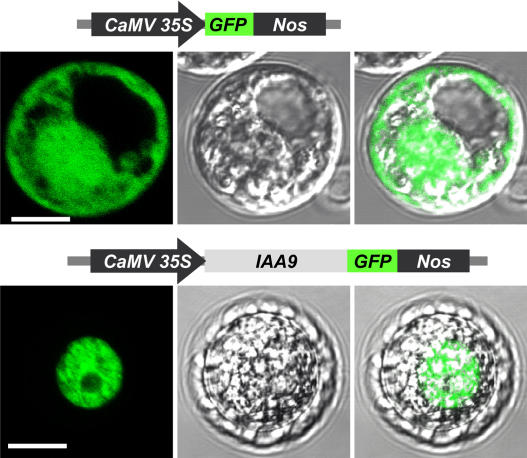
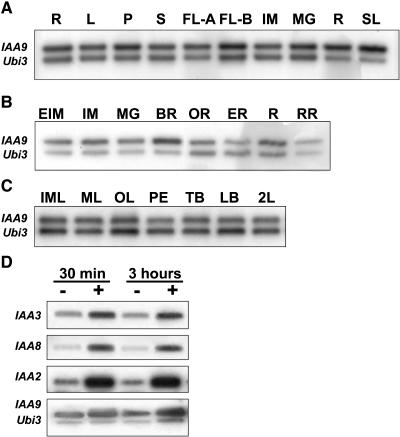
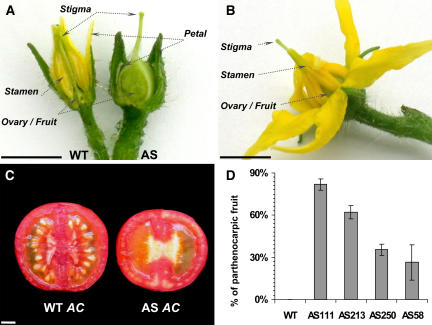
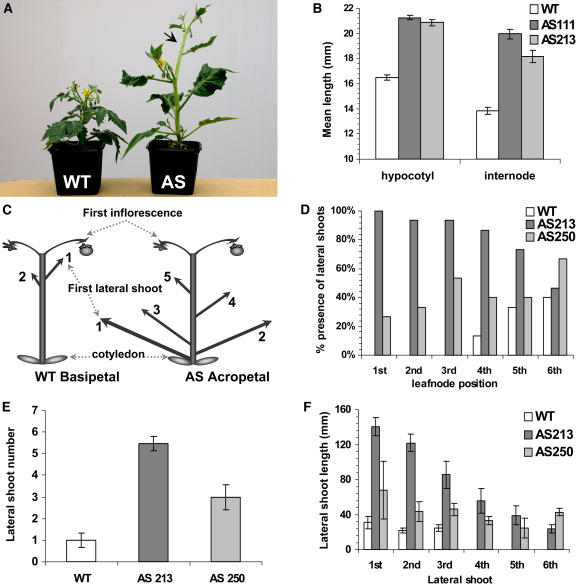
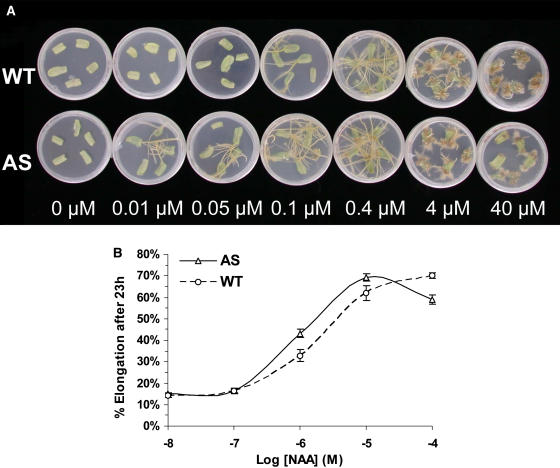
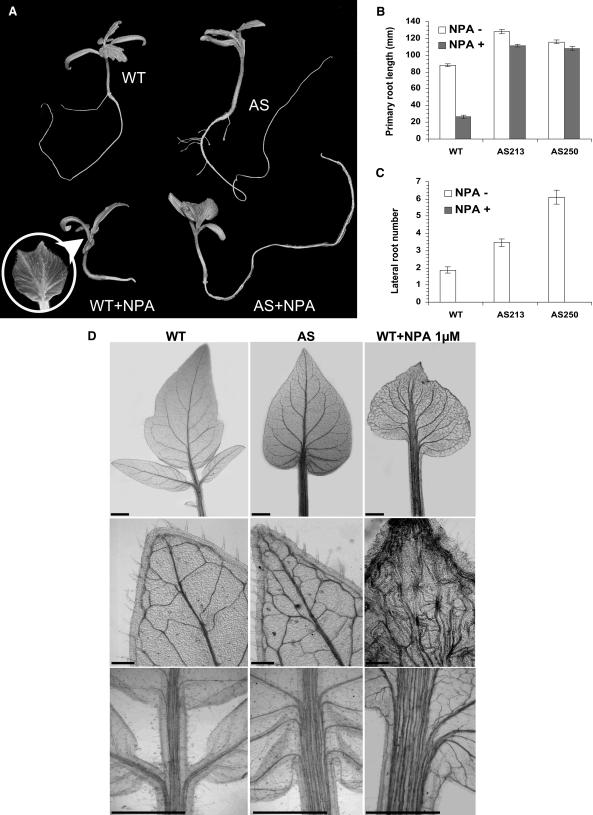
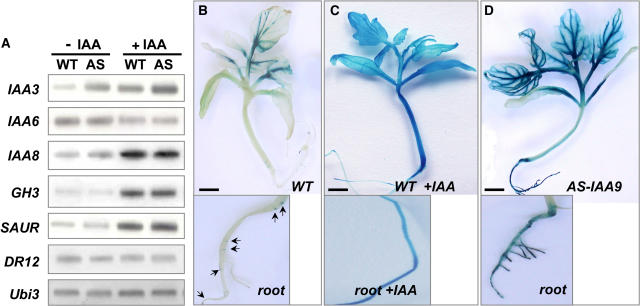
Auxin/indole-3-acetic acid (Aux/IAA) proteins are transcriptional regulators that mediate many aspects of plant responses to auxin. While functions of most Aux/IAAs have been defined mainly by gain-of-function mutant alleles in Arabidopsis thaliana, phenotypes associated with loss-of-function mutations have been scarce and subtle. We report here that the downregulation of IAA9, a tomato (Solanum lycopersicum) gene from a distinct subfamily of Aux/IAA genes, results in a pleiotropic phenotype, consistent with its ubiquitous expression pattern. IAA9-inhibited lines have simple leaves instead of wild-type compound leaves, and fruit development is triggered before fertilization, giving rise to parthenocarpy. This indicates that IAA9 is a key mediator of leaf morphogenesis and fruit set. In addition, antisense plants displayed auxin-related growth alterations, including enhanced hypocotyl/stem elongation, increased leaf vascularization, and reduced apical dominance. Auxin dose-response assays revealed that IAA9 downregulated lines were hypersensitive to auxin, although the only early auxin-responsive gene that was found to be upregulated in the antisense lines was IAA3. The activity of the IAA3 promoter was stimulated in the IAA9 antisense genetic background, indicating that IAA9 acts in planta as a transcriptional repressor of auxin signaling. While no mutation in any member of subfamily IV has been reported to date, the phenotypes associated with the downregulation of IAA9 reveal distinct and novel roles for members of the Aux/IAA gene family.
Figures


References
-
- Abel, S., Nguyen, M.D., and Theologis, A. (1995). The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana. J. Mol. Biol. 251, 533–549. - PubMed
-
- Abel, S., and Theologis, A. (1994). Transient transformation of Arabidopsis leaf protoplasts: A versatile experimental system to study gene expression. Plant J. 5, 421–427. - PubMed
-
- Ainley, W.M., Walker, J.C., Nagao, R.T., and Key, J.L. (1988). Sequence and characterization of two auxin-regulated genes from soybean. J. Biol. Chem. 263, 10658–10666. - PubMed
-
- Beyer, E.M., and Quebedeaux, B. (1974). Parthenocarpy in cucumber: Mechanism of action of auxin transport inhibitors. J. Am. Soc. Hortic. Sci. 99, 385–390.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
