

CXCR 3 activation promotes lymphocyte transendothelial migration across human hepatic endothelium under fluid flow
- PMID: 16127166
- PMCID: PMC1698725
- DOI: 10.1016/S0002-9440(10)62060-3
CXCR 3 activation promotes lymphocyte transendothelial migration across human hepatic endothelium under fluid flow
Abstract
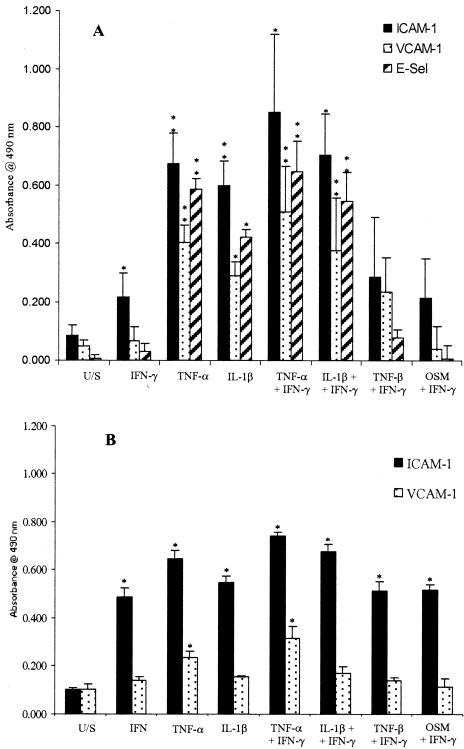
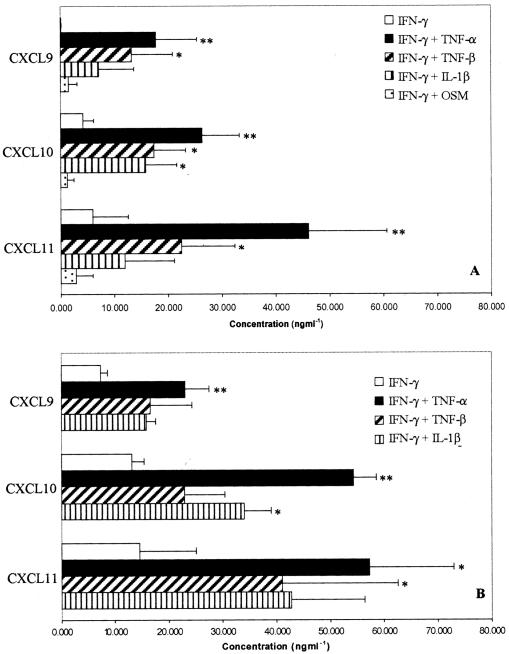
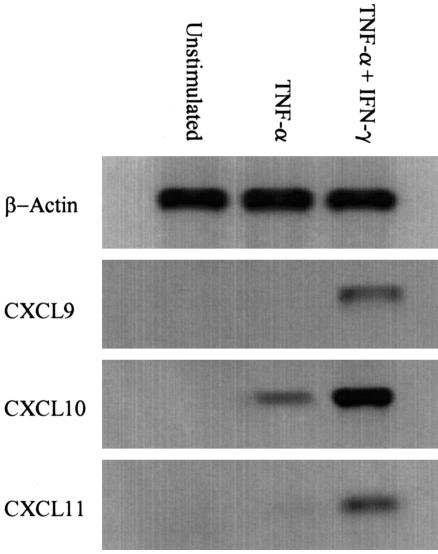
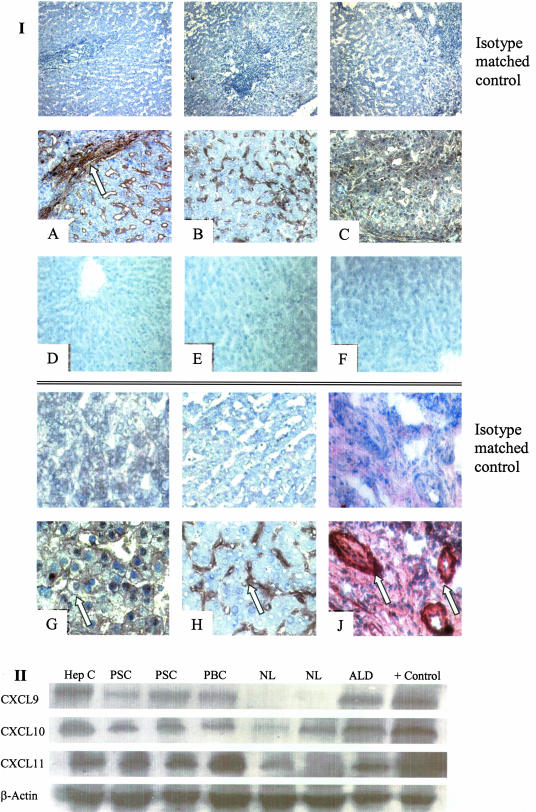
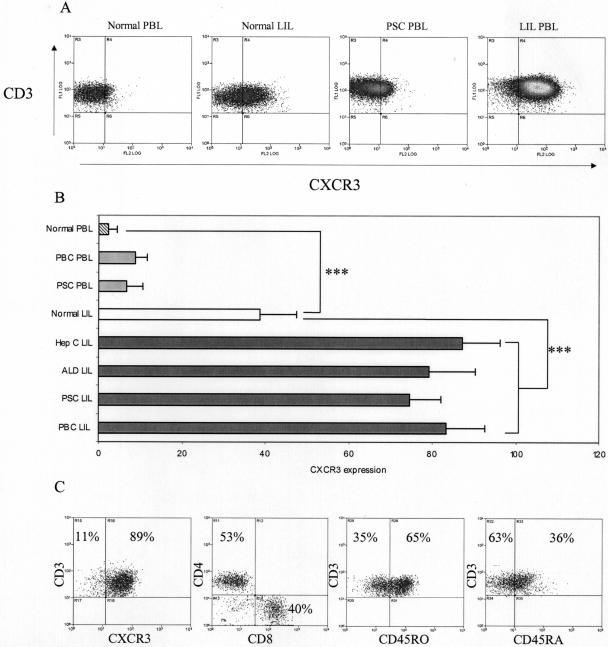
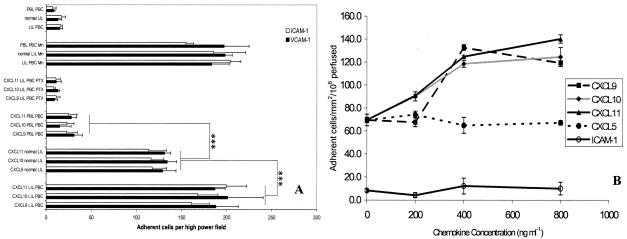
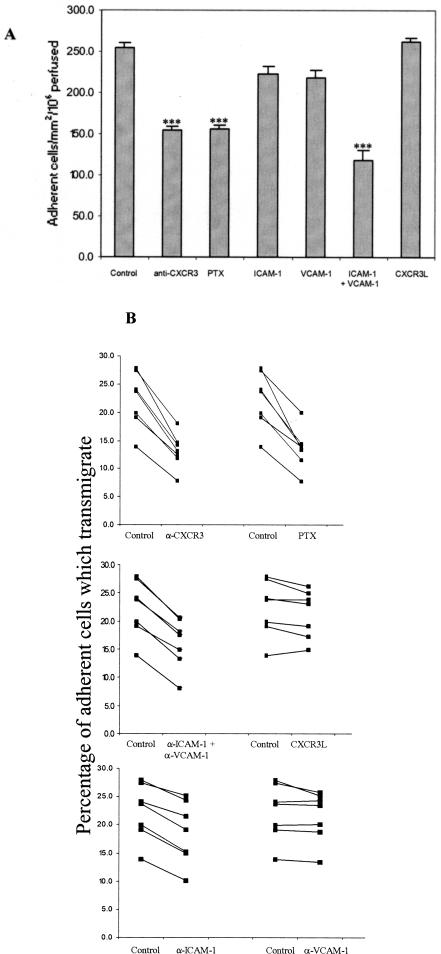
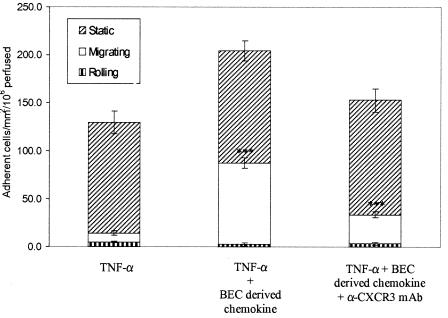
T cells infiltrating the inflamed liver express high levels of CXCR 3 and show enhanced migration to CXCR 3 ligands in chemotactic assays. Moreover, CXCR 3 ligands are up-regulated on hepatic endothelium at sites of T-cell infiltration in chronic hepatitis, and their presence correlates with outcome of inflammatory liver disease. We used a flow-based adhesion assay with human hepatic endothelium to investigate the function of CXCR 3 on lymphocyte adhesion to and transmigration through hepatic endothelium under physiological conditions of blood flow. To more accurately model the function of in vivo activated CXCR 3(high) lymphocytes, we isolated T cells from human liver tissue and studied their behavior in flow-based adhesion assays. We demonstrate that CXCR 3 not only promoted the adhesion of effector T cells to endothelium from flow but also drove transendothelial migration. Moreover, these responses could be stimulated either by endogenous CXCR 3 ligands secreted by the endothelium or by exogenous CXCR 3 ligands derived from other cell types and presented by the endothelium. This study thus demonstrates that activation of CXCR 3 promotes lymphocyte adhesion and transendothelial migration under flow and that human hepatic endothelium can present functionally active chemokines secreted by other cell types within the liver.
Figures
References
-
- Campbell JJ, Butcher EC. Chemokines in tissue-specific and microenvironment-specific lymphocyte homing. Curr Opin Immunol. 2000;12:336–341. - PubMed
-
- von Andrian UH, Mackay CR. T-cell function and migration. Two sides of the same coin. N Engl J Med. 2000;343:1020–1034. - PubMed
-
- Pachynski RK, Wu SW, Gunn MD, Erle DJ. Secondary lymphoid-tissue chemokine (SLC) stimulates integrin alpha 4 beta 7-mediated adhesion of lymphocytes to mucosal addressin cell adhesion molecule-1 (MAdCAM-1) under flow. J Immunol. 1998;161:952–956. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
