

Subunit interaction determines IKs participation in cardiac repolarization and repolarization reserve
- PMID: 16129795
- PMCID: PMC1820744
- DOI: 10.1161/CIRCULATIONAHA.105.543306
Subunit interaction determines IKs participation in cardiac repolarization and repolarization reserve
Erratum in
- Circulation. 2009 Sep 8;120(10):e84
Abstract
Background: The role of IKs, the slow delayed rectifier K+ current, in cardiac ventricular repolarization has been a subject of debate.
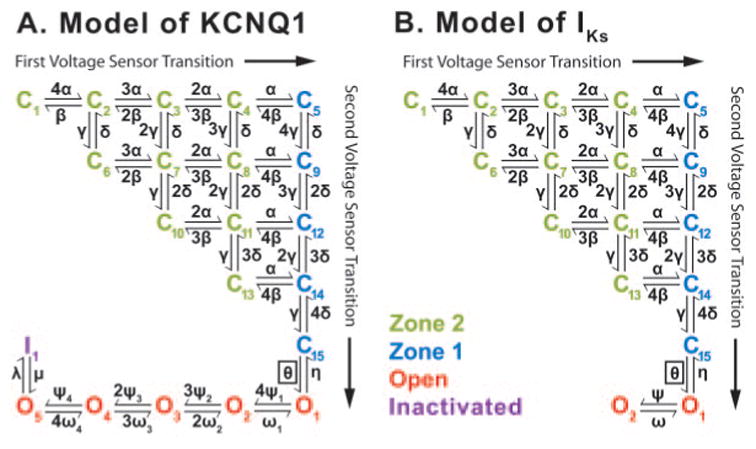
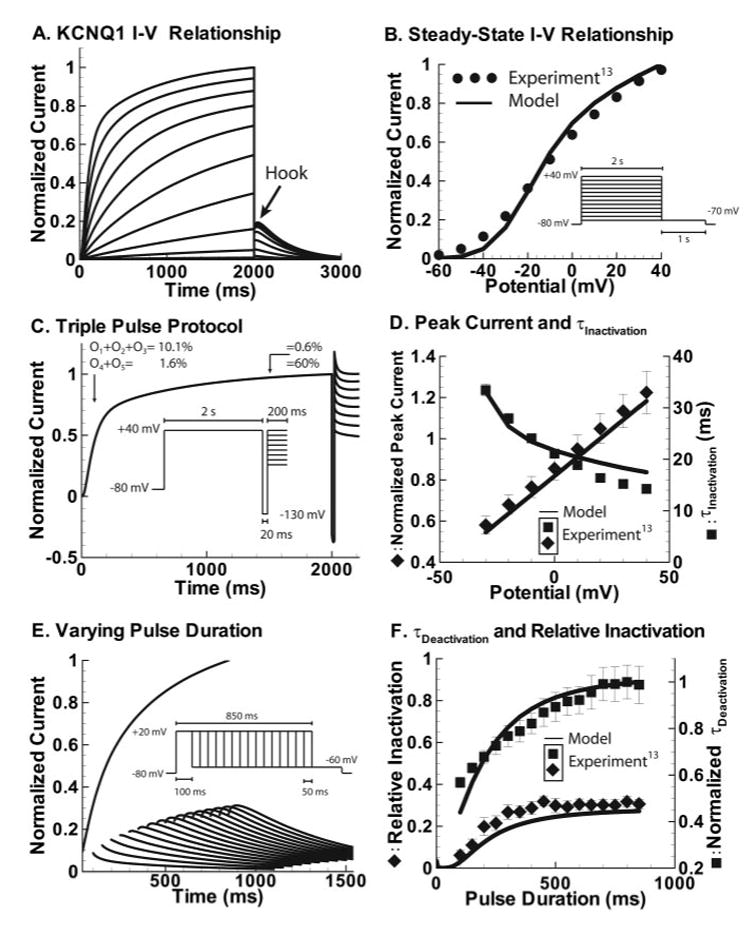
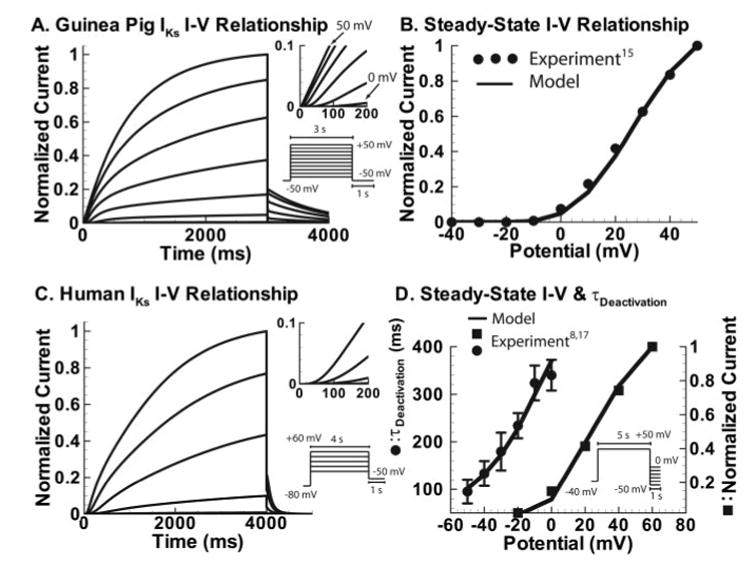
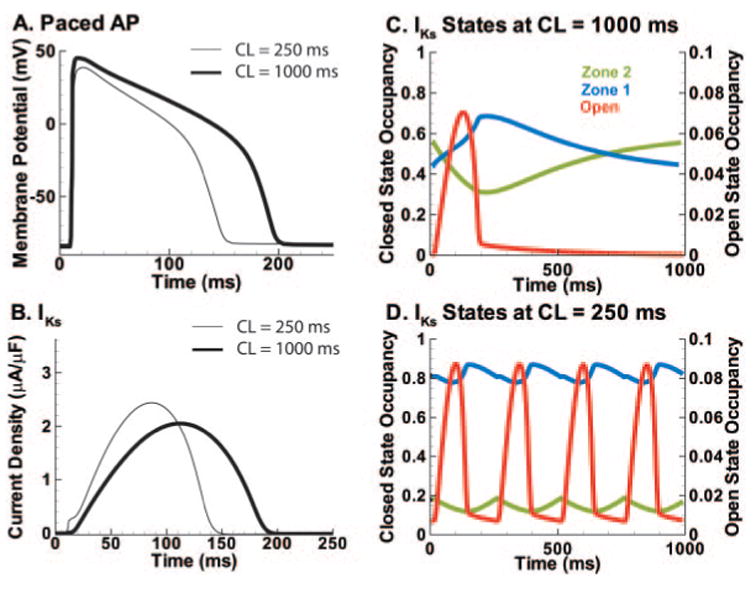
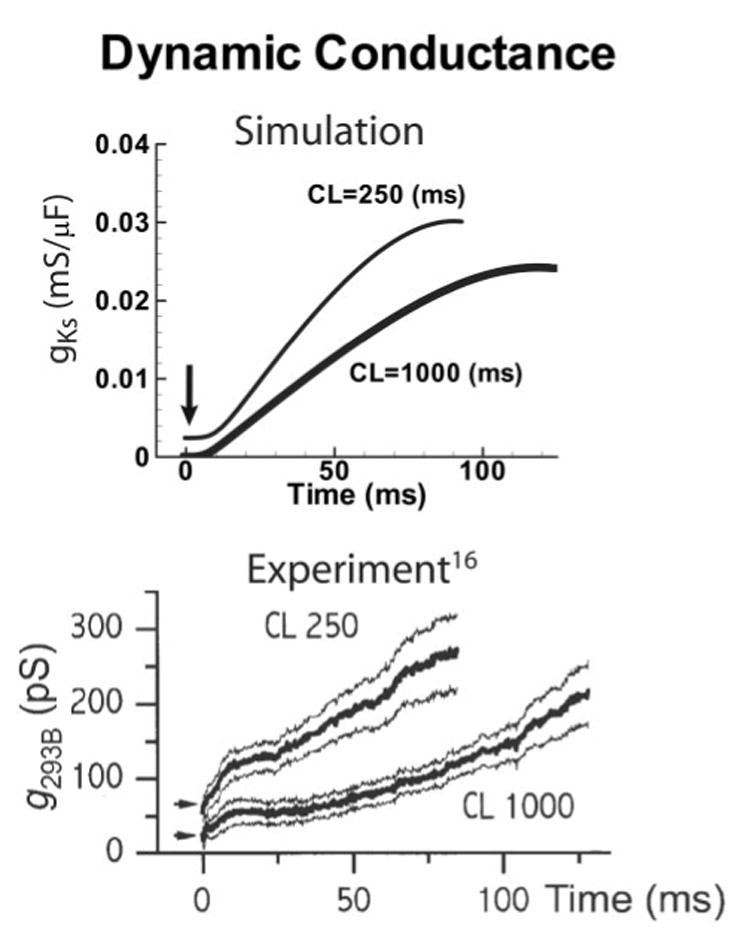
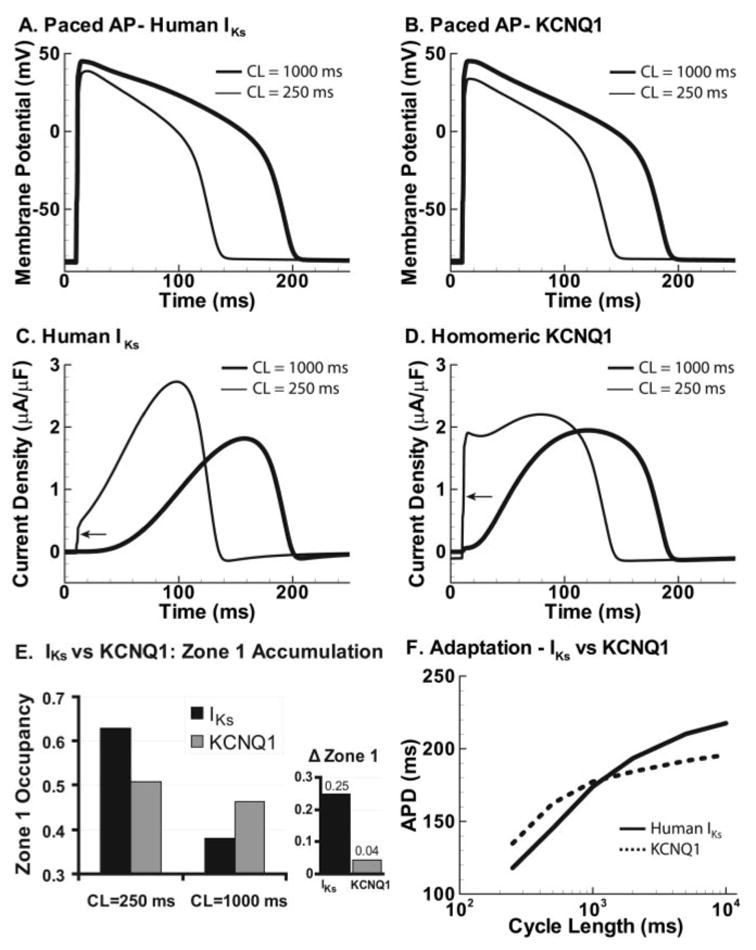
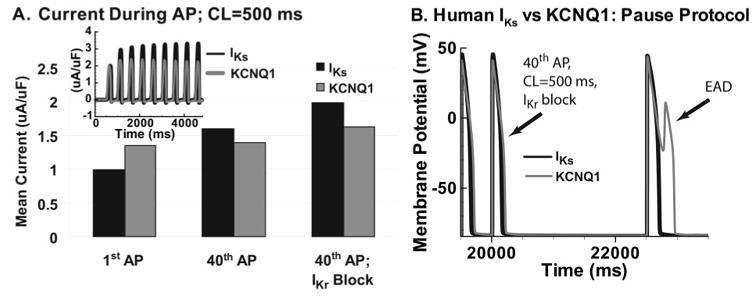
Methods and results: We develop a detailed Markov model of IKs and its alpha-subunit KCNQ1 and examine their kinetic properties during the cardiac ventricular action potential at different rates. We observe that interaction between KCNQ1 and KCNE1 (the beta-subunit) confers kinetic properties on IKs that make it suitable for participation in action potential repolarization and its adaptation to rate changes; in particular, the channel develops an available reserve of closed states near the open state that can open rapidly on demand.
Conclusions: Because of its ability to form an available reserve, IKs can function as a repolarization reserve when IKr, the rapid delayed rectifier, is reduced by disease or drug and can prevent excessive action potential prolongation and development of arrhythmogenic early afterdepolarizations.
Figures
Comment in
-
Protecting the heart against arrhythmias: potassium current physiology and repolarization reserve.Circulation. 2005 Sep 6;112(10):1376-8. doi: 10.1161/CIRCULATIONAHA.105.562777. Circulation. 2005. PMID: 16145010 No abstract available.
References
-
- Sanguinetti MC, Curran ME, Zou A, Shen J, Spector PS, Atkinson DL, Keating MT. Coassembly of K(V)LQT1 and minK (IsK) proteins to form cardiac I(Ks) potassium channel. Nature. 1996;384:80–83. - PubMed
-
- Splawski I, Tristani-Firouzi M, Lehmann MH, Sanguinetti MC, Keating MT. Mutations in the hminK gene cause long QT syndrome and suppress IKs function. Nat Genet. 1997;17:338–340. - PubMed
-
- Antzelevitch C, Dumaine R. Electrical heterogeneity in the heart: physiological, pharmacological and clinical implications. In: Page E, Fozzard H, Solaro J, editors. Handbook of Physiology, Section 2: The Cardiovascular System, Volume I: The Heart. New York, NY: Oxford University Press; 2002. pp. 654–692.
-
- Marx SO, Kurokawa J, Reiken S, Motoike H, D’Armiento J, Marks AR, Kass RS. Requirement of a macromolecular signaling complex for beta adrenergic receptor modulation of the KCNQ1-KCNE1 potassium channel. Science. 2002;295:496–499. - PubMed
-
- Sicouri S, Quist M, Antzelevitch C. Evidence for the presence of M cells in the guinea pig ventricle. J Cardiovasc Electrophysiol. 1996;7:503–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
