

DICER-LIKE 4 functions in trans-acting small interfering RNA biogenesis and vegetative phase change in Arabidopsis thaliana
- PMID: 16129836
- PMCID: PMC1200315
- DOI: 10.1073/pnas.0506426102
DICER-LIKE 4 functions in trans-acting small interfering RNA biogenesis and vegetative phase change in Arabidopsis thaliana
Abstract
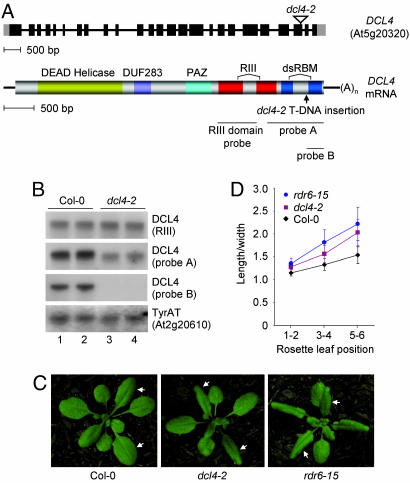
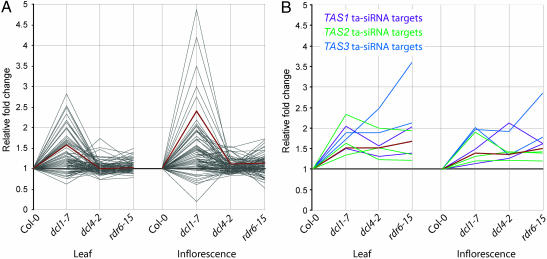
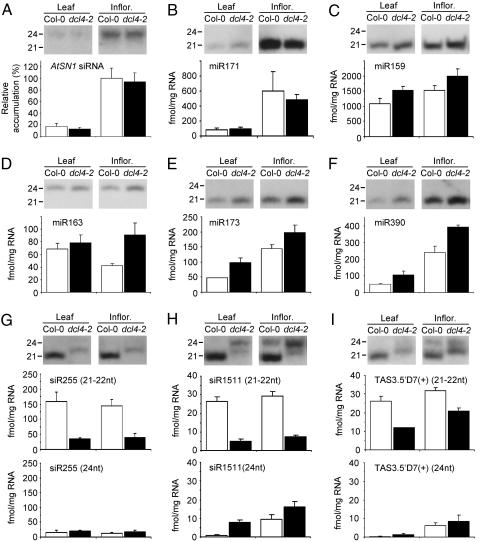
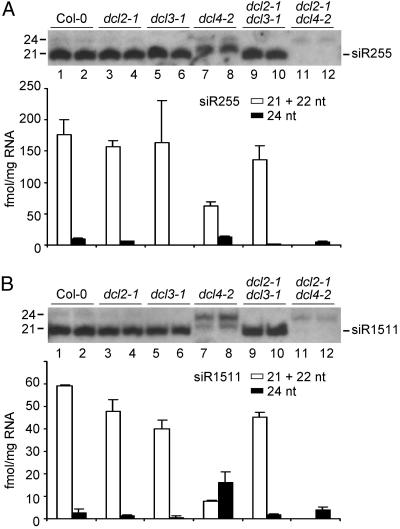
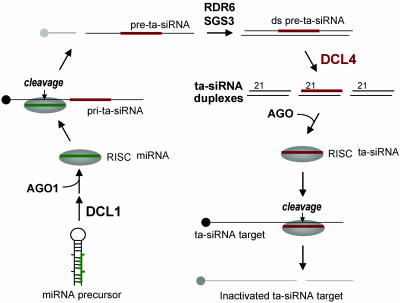
Arabidopsis thaliana contains four DICER-LIKE (DCL) genes with specialized functions in small RNA biogenesis for RNA interference-related processes. A mutant with defects in DCL4 was identified and analyzed for microRNA- and endogenous, small interfering RNA (siRNA)-related functions. The dcl4-2 mutant contained normal or near-normal levels of microRNAs (21 nt) and heterochromatin-associated siRNAs (24 nt). In contrast, this mutant lacked each of three families of 21-nt trans-acting siRNAs (ta-siRNAs) and possessed elevated levels of ta-siRNA target transcripts. The dcl4-2 mutant resembled an rna-dependent RNA polymerase 6 mutant in that both mutants lacked ta-siRNAs and displayed heterochronic defects in which vegetative phase change was accelerated. Double mutant analyses with dcl2-1, dcl3-1, and dcl4-2 alleles revealed hierarchical redundancy among DCL activities, leading to alternative processing of ta-siRNA precursors in the absence of DCL4. These data support the concept that plants have specialized and compartmentalized DCL functions for biogenesis of distinct small RNA classes.
Figures
References
-
- Finnegan, E. J. & Matzke, M. A. (2003) J. Cell Sci. 116, 4689-4693. - PubMed
-
- Bartel, D. (2004) Cell 116, 281-297. - PubMed
-
- Denli, A. M. & Hannon, G. J. (2003) Trends Biochem. Sci. 28, 196-201. - PubMed
-
- Baulcombe, D. (2004) Nature 431, 356-363. - PubMed
-
- Khvorova, A., Reynolds, A. & Jayasena, S. D. (2003) Cell 115, 209-216. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
