

Comparative and evolutionary analysis of the bacterial homologous recombination systems
- PMID: 16132081
- PMCID: PMC1193525
- DOI: 10.1371/journal.pgen.0010015
Comparative and evolutionary analysis of the bacterial homologous recombination systems
Abstract
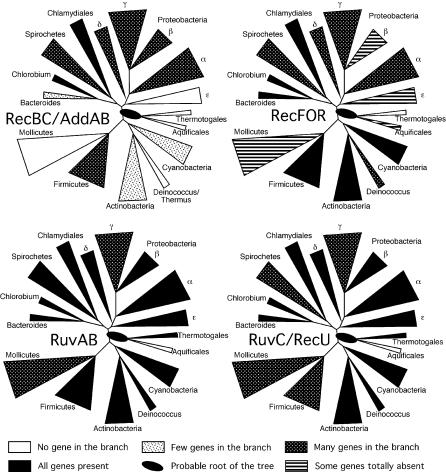
Homologous recombination is a housekeeping process involved in the maintenance of chromosome integrity and generation of genetic variability. Although detailed biochemical studies have described the mechanism of action of its components in model organisms, there is no recent extensive assessment of this knowledge, using comparative genomics and taking advantage of available experimental data on recombination. Using comparative genomics, we assessed the diversity of recombination processes among bacteria, and simulations suggest that we missed very few homologs. The work included the identification of orthologs and the analysis of their evolutionary history and genomic context. Some genes, for proteins such as RecA, the resolvases, and RecR, were found to be nearly ubiquitous, suggesting that the large majority of bacterial genomes are capable of homologous recombination. Yet many genomes show incomplete sets of presynaptic systems, with RecFOR being more frequent than RecBCD/AddAB. There is a significant pattern of co-occurrence between these systems and antirecombinant proteins such as the ones of mismatch repair and SbcB, but no significant association with nonhomologous end joining, which seems rare in bacteria. Surprisingly, a large number of genomes in which homologous recombination has been reported lack many of the enzymes involved in the presynaptic systems. The lack of obvious correlation between the presence of characterized presynaptic genes and experimental data on the frequency of recombination suggests the existence of still-unknown presynaptic mechanisms in bacteria. It also indicates that, at the moment, the assessment of the intrinsic stability or recombination isolation of bacteria in most cases cannot be inferred from the identification of known recombination proteins in the genomes.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures





References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
