

Pocket protein complexes are recruited to distinct targets in quiescent and proliferating cells
- PMID: 16135806
- PMCID: PMC1234327
- DOI: 10.1128/MCB.25.18.8166-8178.2005
Pocket protein complexes are recruited to distinct targets in quiescent and proliferating cells
Abstract
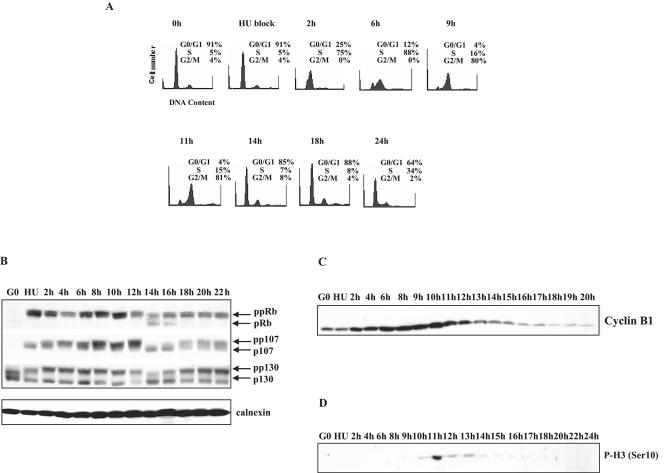
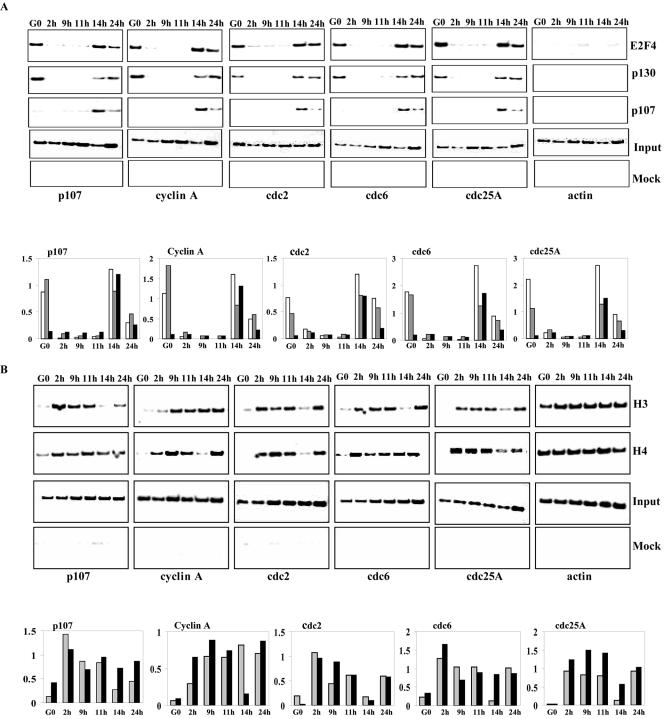
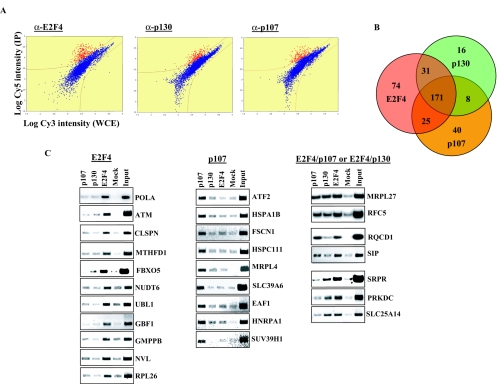
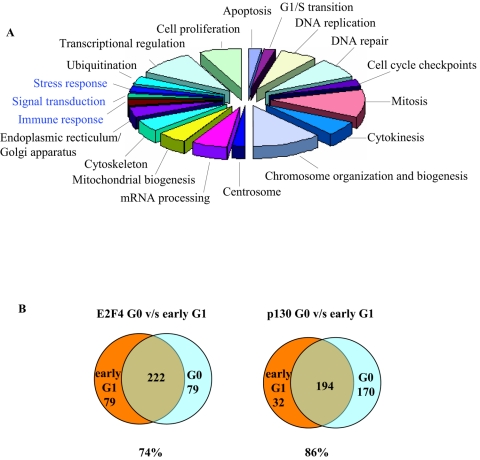
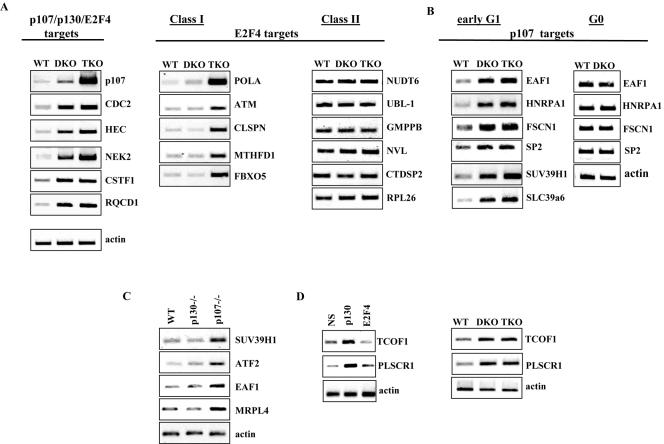
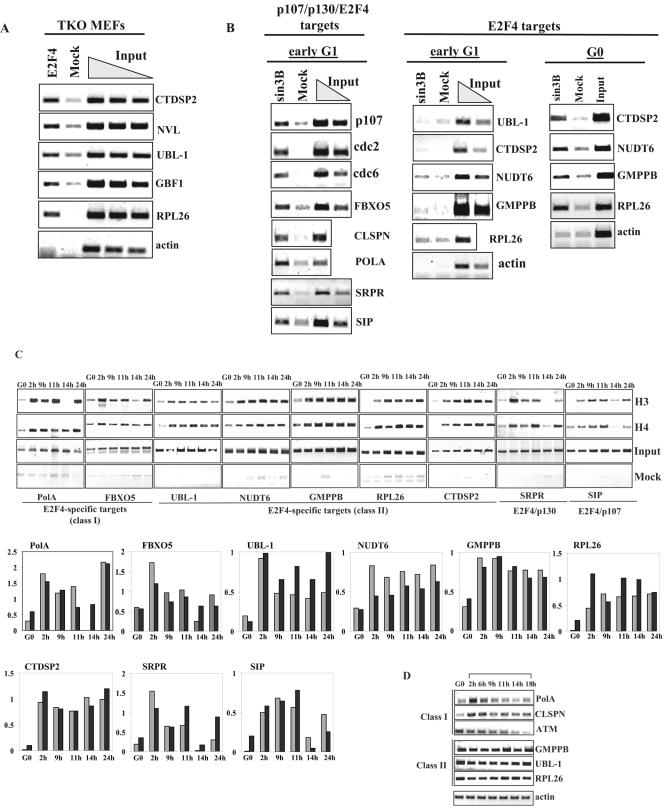
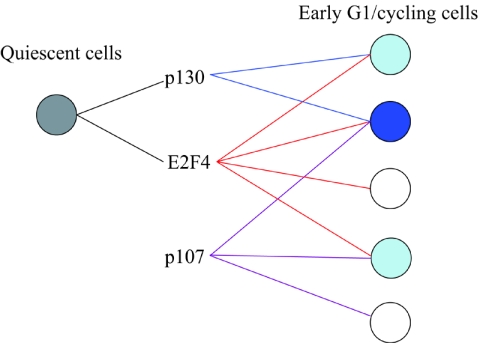
Biochemical and genetic studies have determined that retinoblastoma protein (pRB) tumor suppressor family members have overlapping functions. However, these studies have largely failed to distinguish functional differences between the highly related p107 and p130 proteins. Moreover, most studies pertaining to the pRB family and its principal target, the E2F transcription factor, have focused on cells that have reinitiated a cell cycle from quiescence, although recent studies suggest that cycling cells exhibit layers of regulation distinct from mitogenically stimulated cells. Using genome-wide chromatin immunoprecipitation, we show that there are distinct classes of genes directly regulated by unique combinations of E2F4, p107, and p130, including a group of genes specifically regulated in cycling cells. These groups exhibit both distinct histone acetylation signatures and patterns of mammalian Sin3B corepressor recruitment. Our findings suggest that cell cycle-dependent repression results from recruitment of an unexpected array of diverse complexes and reveals specific differences between transcriptional regulation in cycling and quiescent cells. In addition, factor location analyses have, for the first time, allowed the identification of novel and specific targets of the highly related transcriptional regulators p107 and p130, suggesting new and distinct regulatory networks engaged by each protein in continuously cycling cells.
Figures
References
-
- Alland, L., R. Muhle, H. Hou, Jr., J. Potes, L. Chin, N. Schreiber-Agus, and R. A. DePinho. 1997. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature 387:49-55. - PubMed
-
- Blais, A., and B. D. Dynlacht. 2004. Hitting their targets: an emerging picture of E2F and cell cycle control. Curr. Opin. Genet. Dev. 14:527-532. - PubMed
-
- Cam, H., E. Balciunaite, A. Blais, A. Spektor, R. C. Scarpulla, R. Young, Y. Kluger, and B. D. Dynlacht. 2004. A common set of gene regulatory networks links metabolism and growth inhibition. Mol. Cell 16:399-411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources