

Human subtelomeres are hot spots of interchromosomal recombination and segmental duplication
- PMID: 16136133
- PMCID: PMC1368961
- DOI: 10.1038/nature04029
Human subtelomeres are hot spots of interchromosomal recombination and segmental duplication
Abstract
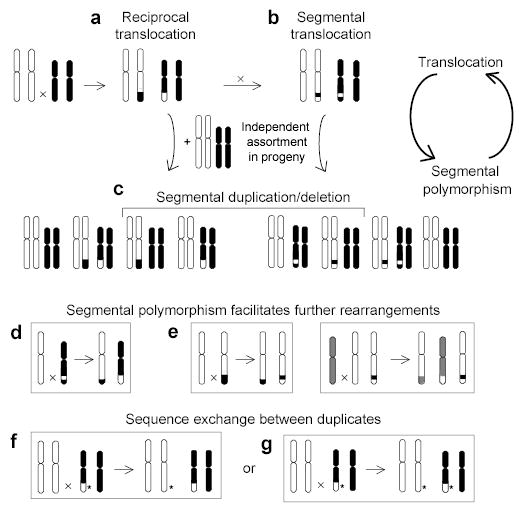
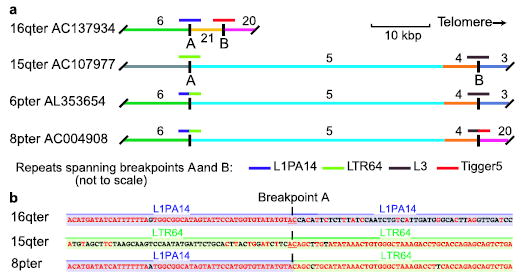
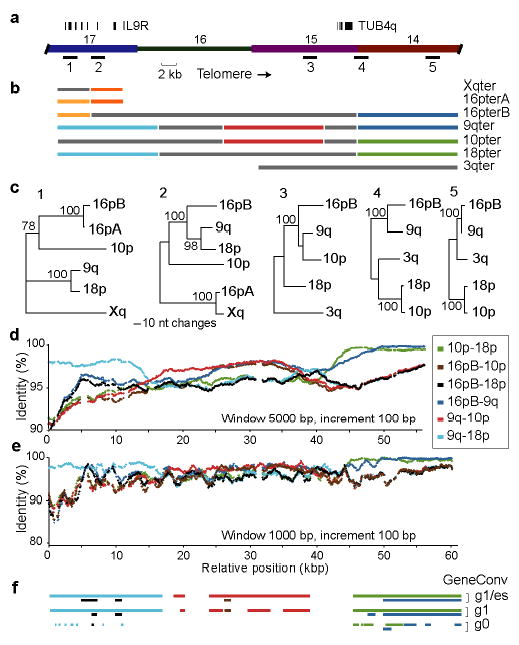
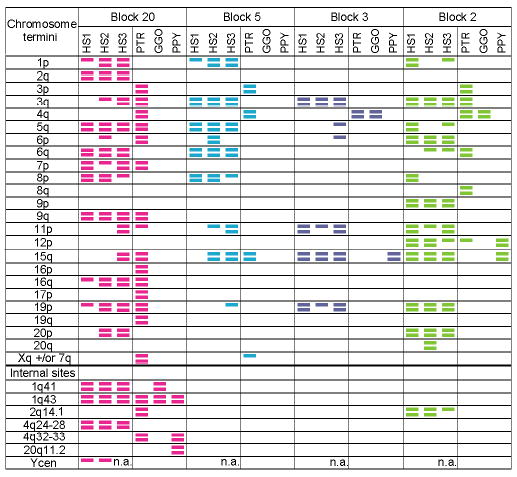
Human subtelomeres are polymorphic patchworks of interchromosomal segmental duplications at the ends of chromosomes. Here we provide evidence that these patchworks arose recently through repeated translocations between chromosome ends. We assess the relative contribution of the principal mechanisms of ectopic DNA repair to the formation of subtelomeric duplications and find that non-homologous end-joining predominates. Once subtelomeric duplications arise, they are prone to homology-based sequence transfers as shown by the incongruent phylogenetic relationships of neighbouring sections. Interchromosomal recombination of subtelomeres is a potent force for recent change. Cytogenetic and sequence analyses reveal that pieces of the subtelomeric patchwork have changed location and copy number with unprecedented frequency during primate evolution. Half of the known subtelomeric sequence has formed recently, through human-specific sequence transfers and duplications. Subtelomeric dynamics result in a gene duplication rate significantly higher than the genome average and could have both advantageous and pathological consequences in human biology. More generally, our analyses suggest an evolutionary cycle between segmental polymorphisms and genome rearrangements.
Figures


References
-
- Samonte RV, Eichler EE. Segmental duplications and the evolution of the primate genome. Nat Rev Genet. 2002;3:65–72. - PubMed
-
- Bailey JA, et al. Recent segmental duplications in the human genome. Science. 2002;297:1003–7. - PubMed
-
- Shaw CJ, Lupski JR. Implications of human genome architecture for rearrangement-based disorders: the genomic basis of disease. Hum Mol Genet. 2004;13(Spec No 1):R57–64. - PubMed
-
- Mefford H, Trask BJ. The complex structure and dynamic evolution of human subtelomeres. Nature Reviews Genetics. 2002;3:91–102. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
