

Stronger control of ATP/ADP by proton leak in pancreatic beta-cells than skeletal muscle mitochondria
- PMID: 16137248
- PMCID: PMC1383673
- DOI: 10.1042/BJ20051280
Stronger control of ATP/ADP by proton leak in pancreatic beta-cells than skeletal muscle mitochondria
Abstract
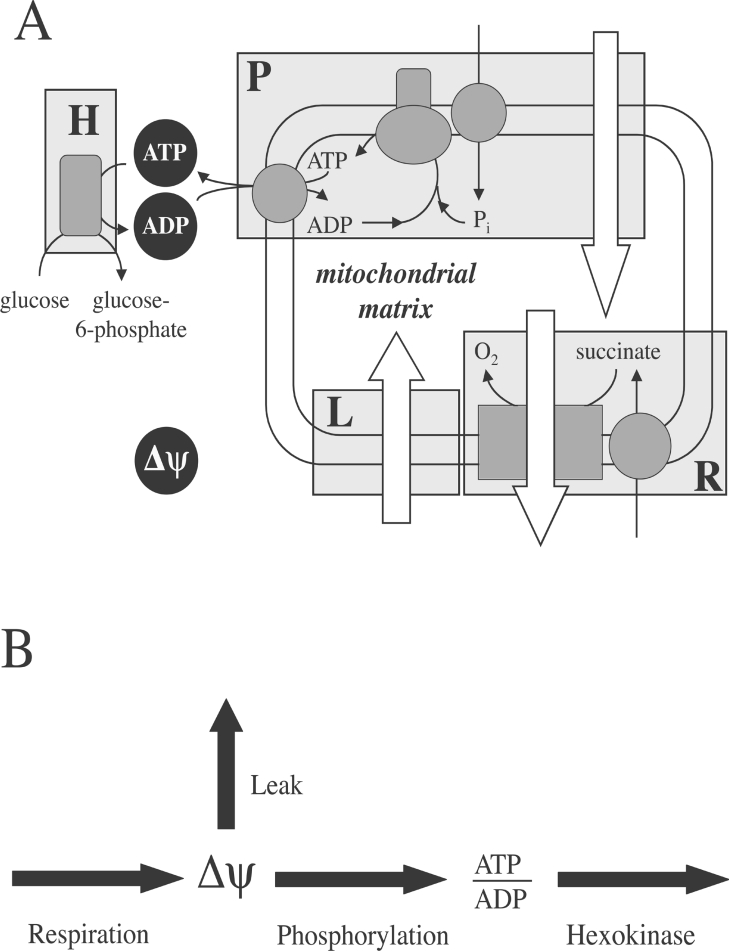
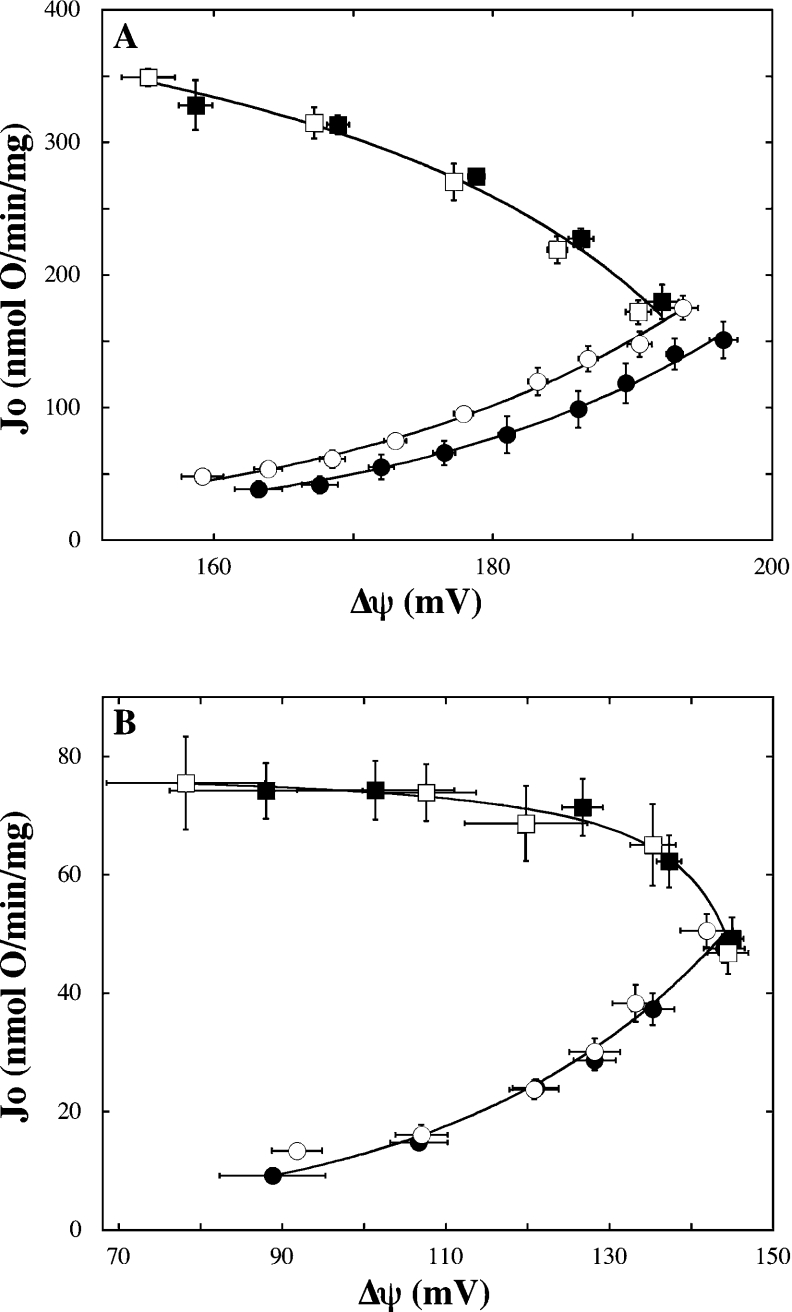
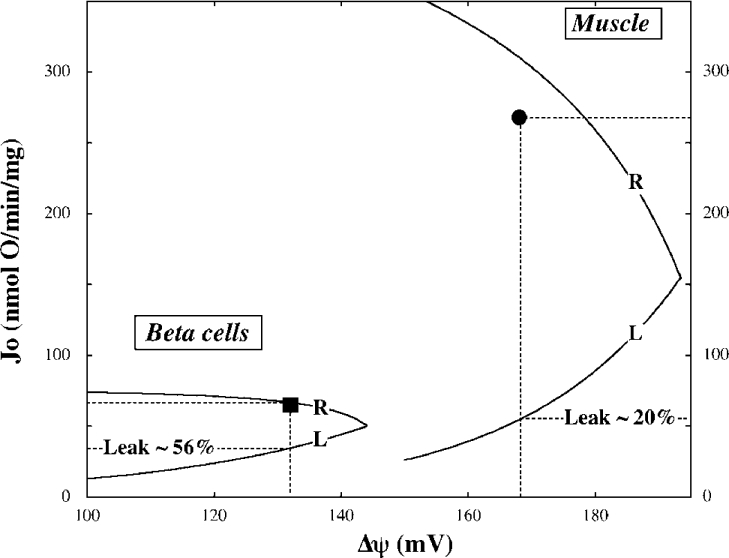
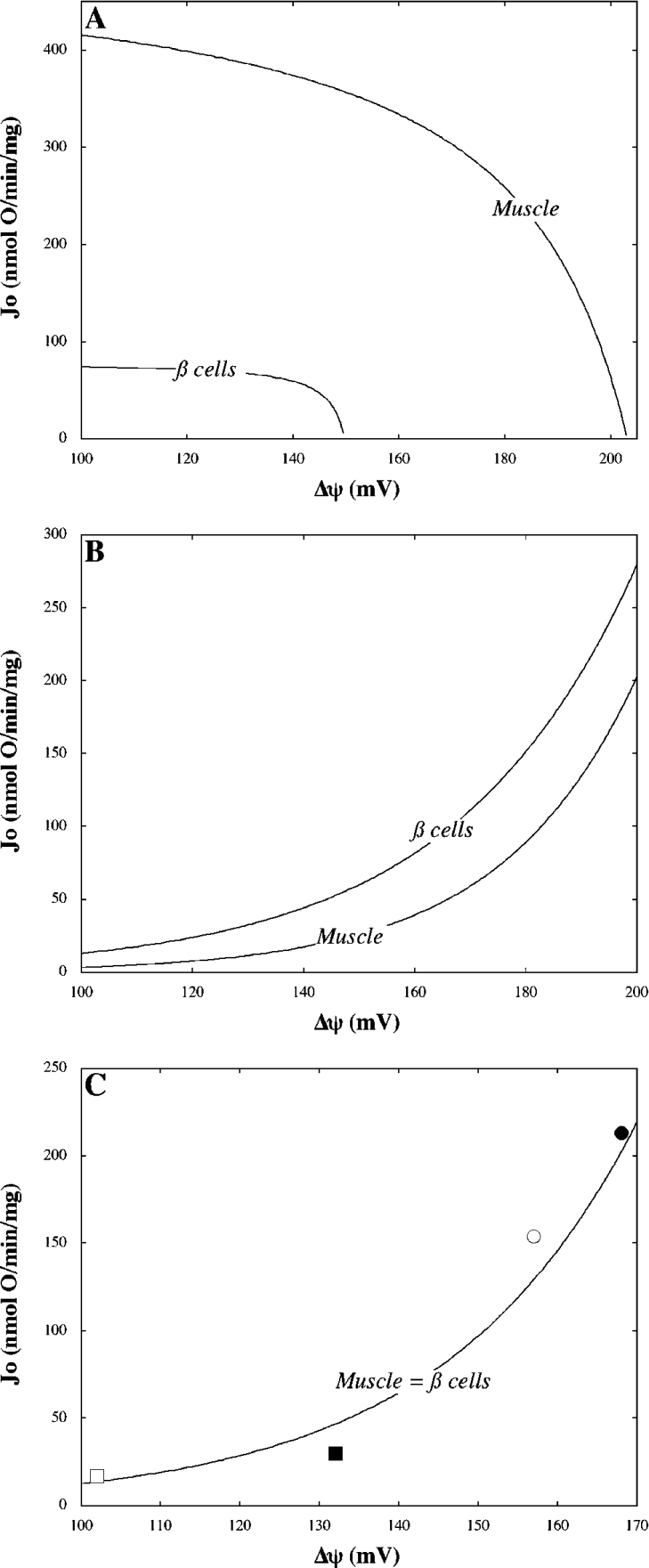
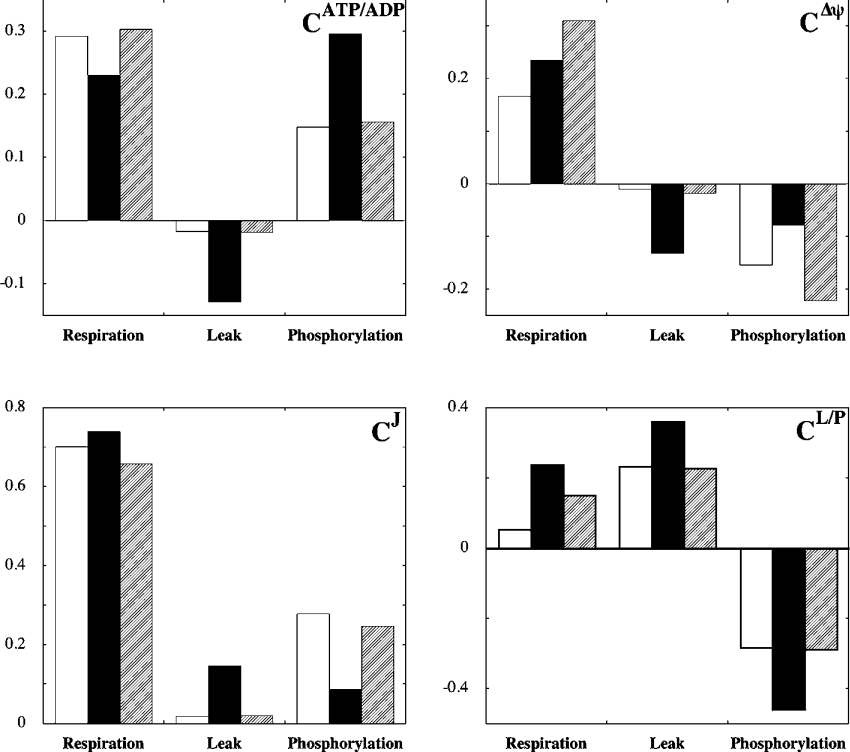
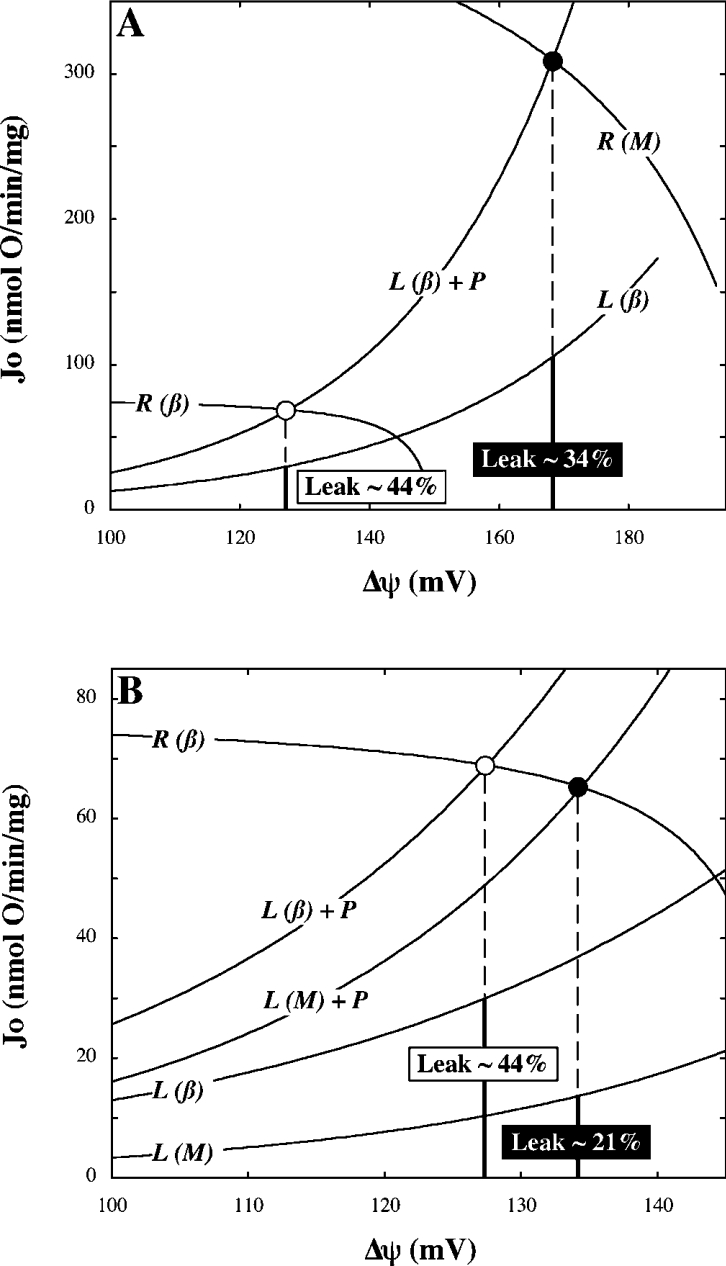
Pancreatic beta cells respond to rising blood glucose concentrations by increasing their oxidative metabolism, which leads to an increased ATP/ADP ratio, closure of K(ATP) channels, depolarization of the plasma membrane potential, influx of calcium and the eventual secretion of insulin. Such a signalling mechanism implies that the ATP/ADP ratio is flexible in beta cells (beta-cells), which is in contrast with other cell types (e.g. muscle and liver) that maintain a stable ATP/ADP poise while respiring at widely varying rates. To determine whether this difference in flexibility is accounted for by mitochondrial peculiarities, we performed a top-down metabolic control analysis to quantitatively assess how ATP/ADP is controlled in mitochondria isolated from rat skeletal muscle and cultured beta cells. We show that the ATP/ADP ratio is more strongly controlled (approx. 7.5-fold) by proton leak in beta cells than in muscle. The comparatively high importance of proton leak in beta cell mitochondria (relative to phosphorylation) is evidenced furthermore by its relatively high level of control over membrane potential and overall respiratory activity. Modular-kinetic analysis of oxidative phosphorylation reveals that these control differences can be fully explained by a higher relative leak activity in beta cell mitochondria, which results in a comparatively high contribution of proton leak to the overall respiratory activity in this system.
Figures
References
-
- Kiberstis P. A. A surfeit of suspects. Science (Washington DC) 2005;307:369.
-
- Lowell B. B., Shulman G. I. Mitochondrial dysfunction and type 2 diabetes. Science (Washington DC) 2005;307:384–387. - PubMed
-
- Rutter G. A. Nutrient-secretion coupling in the pancreatic islet β-cell: recent advances. Mol. Asp. Med. 2001;22:247–284. - PubMed
-
- Ainscow E. K., Brand M. D. Top-down control analysis of ATP turnover, glycolysis and oxidative phosphorylation in rat hepatocytes. Eur. J. Biochem. 1999;263:671–685. - PubMed
-
- Matschinsky F. M., Glaser B., Magnuson M. A. Pancreatic β-cell glucokinase: closing the gap between theoretical concepts and experimental realities. Diabetes. 1998;47:307–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources