

Cytology and functionally correlated circuits of human posterior cingulate areas
- PMID: 16140550
- PMCID: PMC2649771
- DOI: 10.1016/j.neuroimage.2005.07.048
Cytology and functionally correlated circuits of human posterior cingulate areas
Abstract
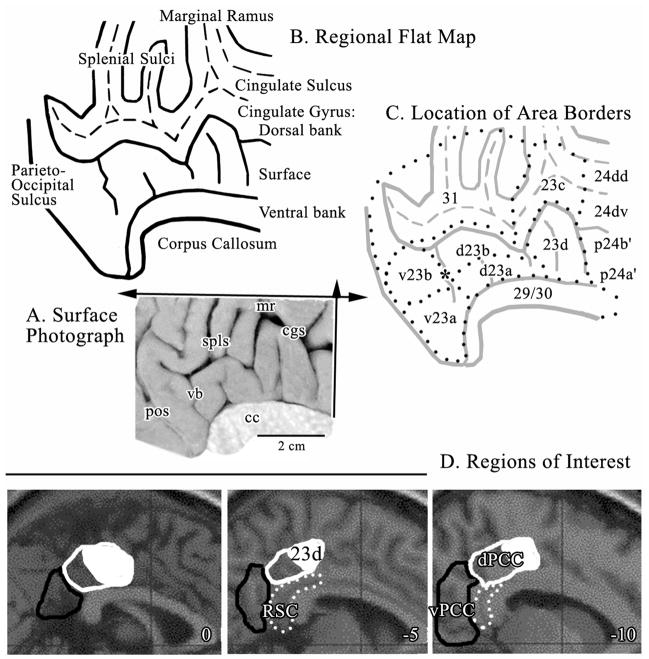
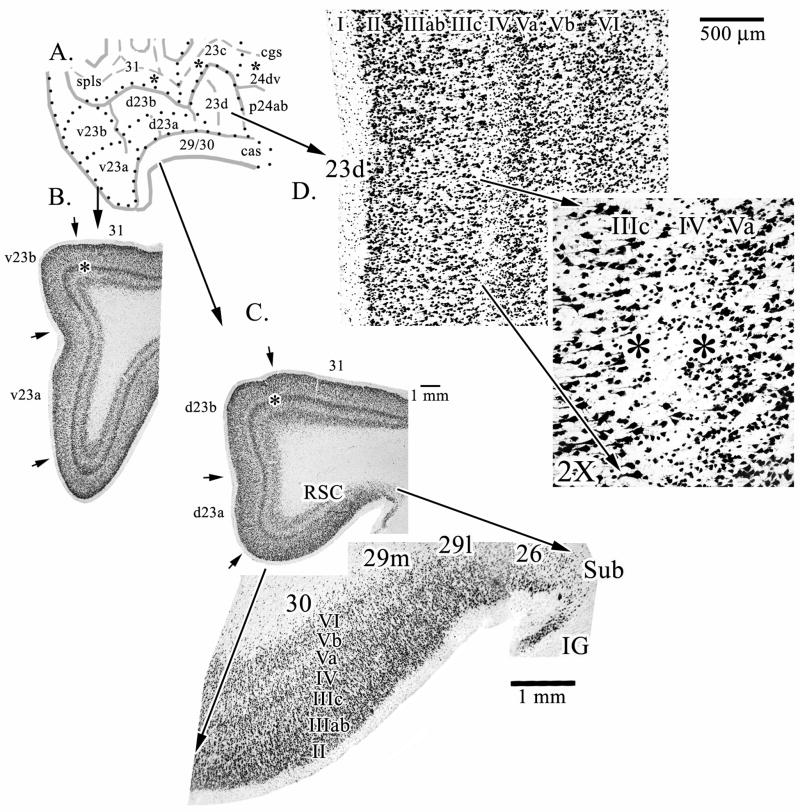
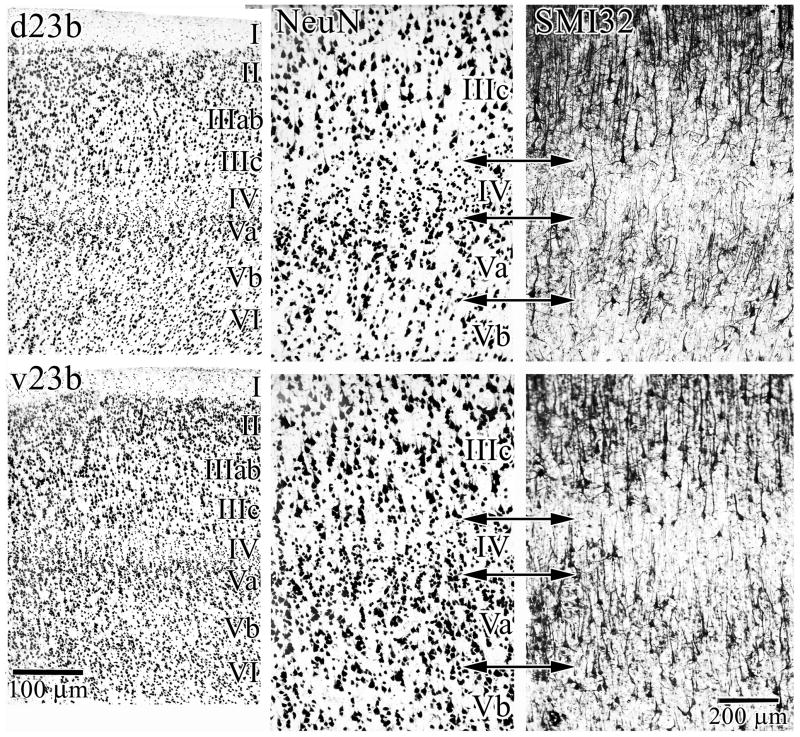
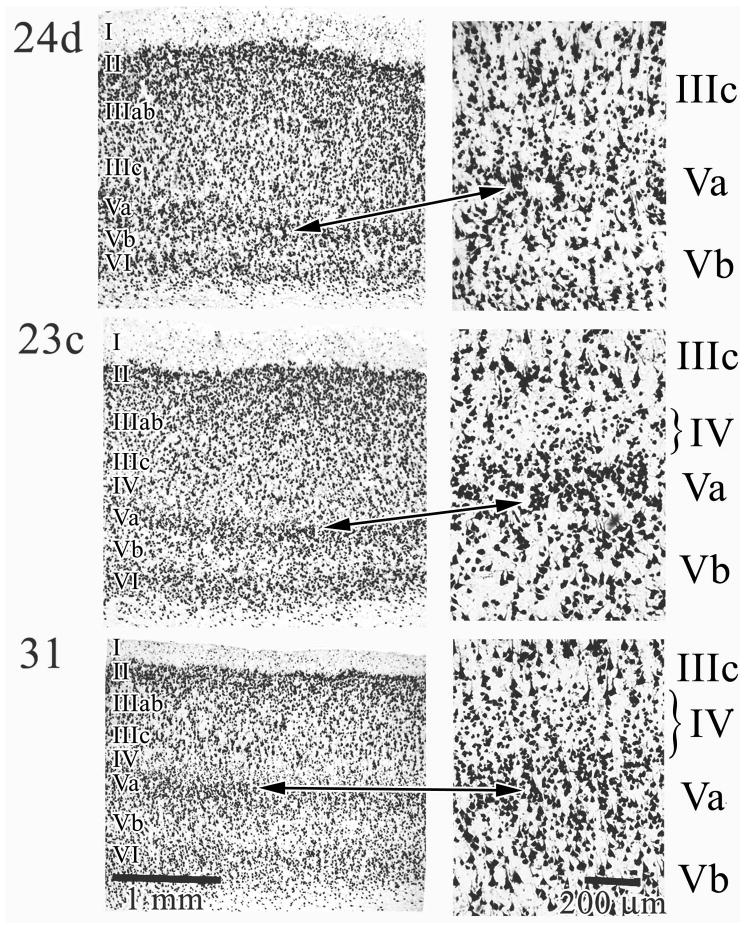
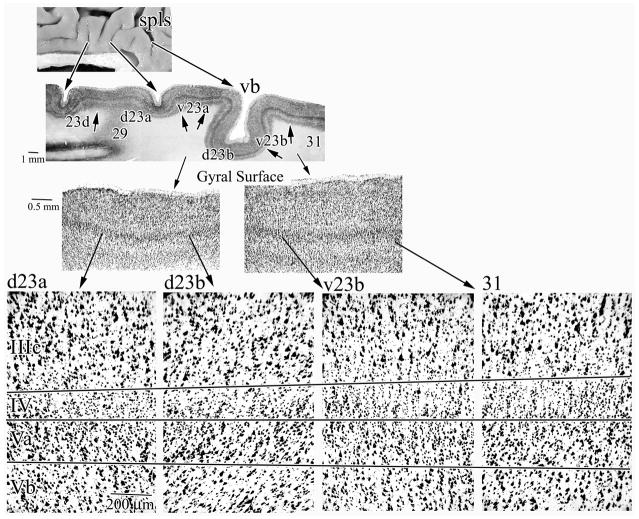
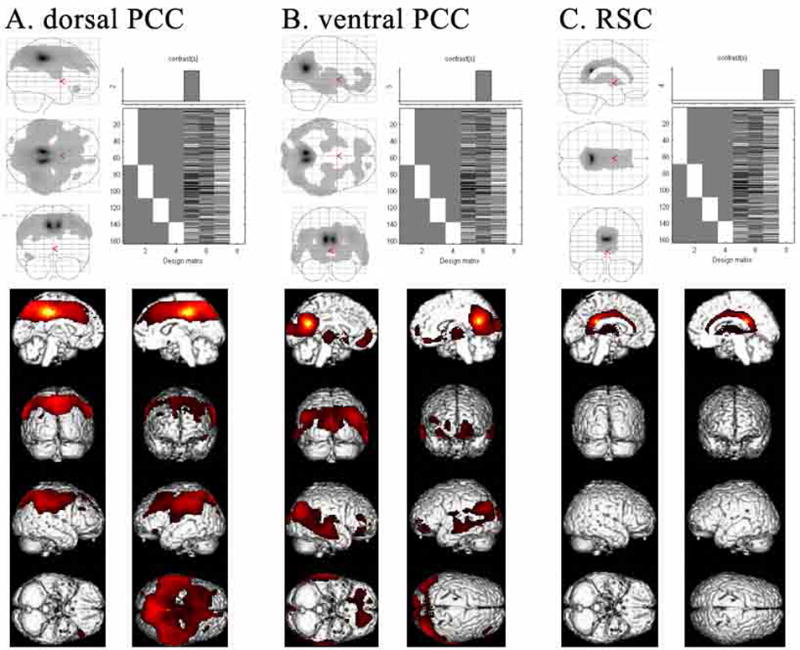
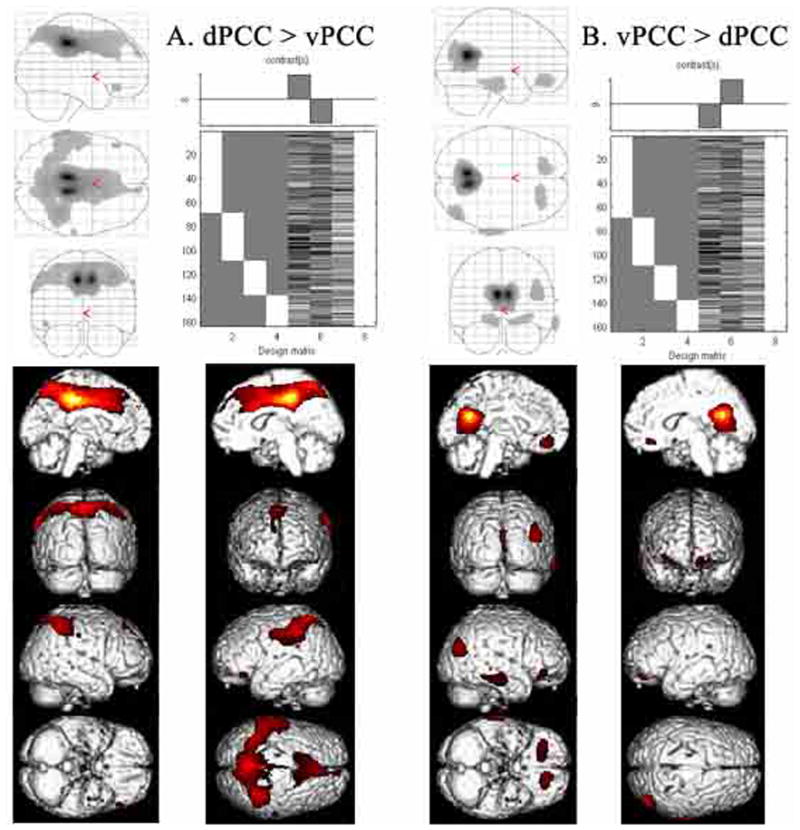
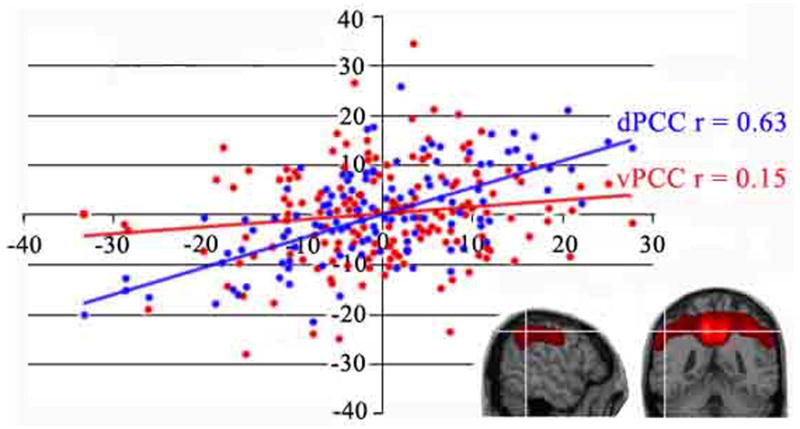
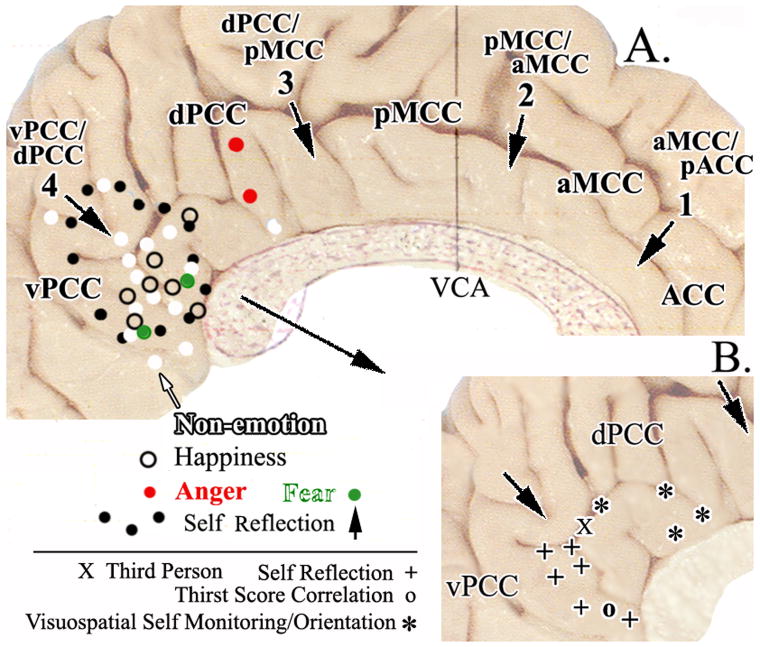
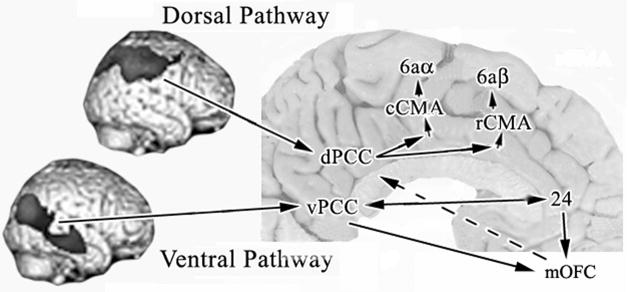
Human posterior cingulate cortex (PCC) and retrosplenial cortex (RSC) form the posterior cingulate gyrus, however, monkey connection and human imaging studies suggest that PCC area 23 is not uniform and atlases mislocate RSC. We histologically assessed these regions in 6 postmortem cases, plotted a flat map, and characterized differences in dorsal (d) and ventral (v) area 23. Subsequently, functional connectivity of histologically guided regions of interest (ROI) were assessed in 163 [(18)F]fluorodeoxyglucose human cases with PET. Compared to area d23, area v23 had a higher density and larger pyramids in layers II, IIIc, and Vb and more intermediate neurofilament-expressing neurons in layer Va. Coregisrtration of each case to standard coordinates showed that the ventral branch of the splenial sulci coincided with the border between d/v PCC at -5.4 +/- 0.17 cm from the vertical plane and +1.97 +/- 0.08 cm from the bi-commissural line. Correlation analysis of glucose metabolism using histologically guided ROIs suggested important circuit differences including dorsal and ventral visual stream inputs, interactions between the vPCC and subgenual cingulate cortex, and preferential relations between dPCC and the cingulate motor region. The RSC, in contrast, had restricted correlated activity with pericallosal cortex and thalamus. Visual information may be processed with an orbitofrontal link for synthesis of signals to drive premotor activity through dPCC. Review of the literature in terms of a PCC duality suggests that interactions of dPCC, including area 23d, orient the body in space via the cingulate motor areas, while vPCC interacts with subgenual cortex to process self-relevant emotional and non-emotional information and objects and self-reflection.
Figures
References
-
- Ashburner J, Andersson JL, Friston KJ. High-dimentional image registration using symmetric priors. NeuroImage. 1999;9:619–628. - PubMed
-
- Bentley DE, Derbyshire SWG, Youell PD, Jones AKP. Caudal cingulate cortex involvement in pain processing: an inter-individual laser evoked potential source localization study using realistic head models. Pain. 2003;102:265–271. - PubMed
-
- Bernstein LJ, Beig S, Siegenthaler AL, Grady CL. The effect of encoding strategy on the neural correlates of memory for faces. Neuropsychologia. 2002;40:86–89. - PubMed
-
- Berthoz A. Hippocampal and parietal contribution to topokinetic and topographic memory. In: Burgess N, Jeffery KJ, O’Keefe J, editors. The Hippocampal and Parietal Foundations of Spatial Cognition. Oxford University Press; Oxford: 1999.
