

Virion instability of human immunodeficiency virus type 1 reverse transcriptase (RT) mutated in the protease cleavage site between RT p51 and the RT RNase H domain
- PMID: 16140771
- PMCID: PMC1212597
- DOI: 10.1128/JVI.79.18.11952-11961.2005
Virion instability of human immunodeficiency virus type 1 reverse transcriptase (RT) mutated in the protease cleavage site between RT p51 and the RT RNase H domain
Abstract
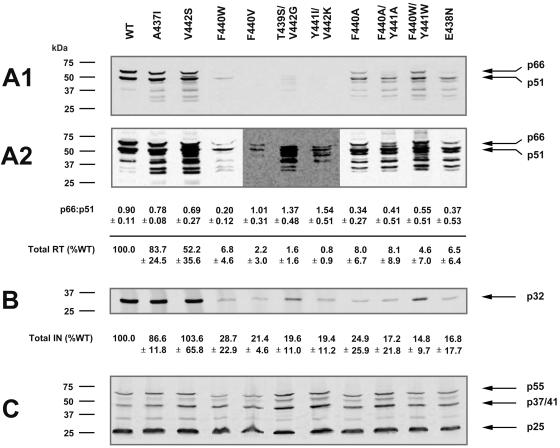
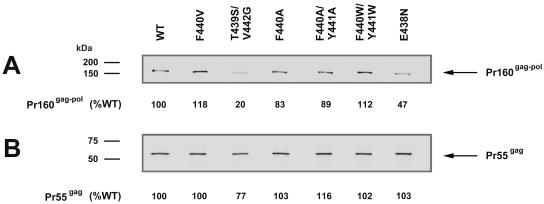
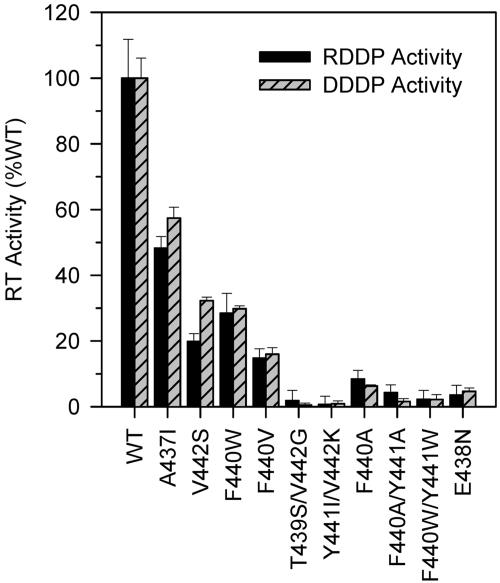
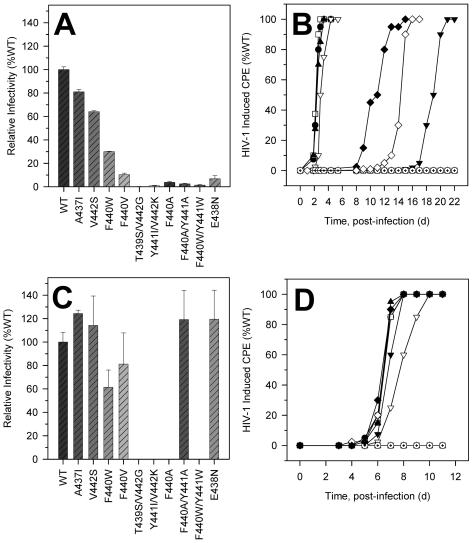
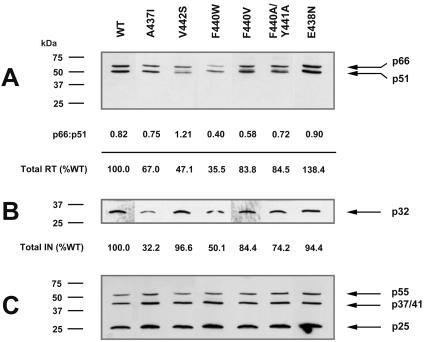
Each of the human immunodeficiency virus type 1 (HIV-1) pol-encoded enzymes, protease (PR), reverse transcriptase (RT), and integrase (IN), is active only as a dimer (or higher-order oligomer in the case of IN), but only RT comprises subunits of different masses. RT is a heterodimer of 66-kDa and 51-kDa subunits. The latter is formed by HIV PR-catalyzed cleavage of p66 during virion maturation, resulting in the removal of the RNase H (RNH) domain of a p66 subunit. In order to study the apparent need for RT heterodimers in the context of the virion, we introduced a variety of mutations in the RT p51-RNH protease cleavage site of an infectious HIV-1 molecular clone. Surprisingly, rather than leading to virions with increased RT p66 content, most of the mutations resulted in significantly attenuated virus that contained greatly decreased levels of RT that in many cases was primarily p51 RT. IN levels were also reduced in several mutants. However, most mutants showed normal levels of the Pr160(gag-pol) precursor polyprotein, suggesting that reduced virion RT arose from proteolytic instability rather than decreased incorporation. Mutant virion p24 Gag levels were equivalent to wild type, indicating that Gag incorporation and processing were not affected. Repeated passage of MT-2 cells exposed to mutant viruses led to the appearance of virus with improved replication capacity; these virions contained normally processed RT at near-wild-type levels. These results imply that additional proteolytic processing of RT to the p66/p51 heterodimer is essential to provide proteolytic stability of RT during HIV-1 maturation.
Figures
References
-
- Amacker, M., and U. Hubscher. 1998. Chimeric HIV-1 and feline immunodeficiency virus reverse transcriptases: critical role of the p51 subunit in the structural integrity of heterodimeric lentiviral DNA polymerases. J. Mol. Biol. 278:757-765. - PubMed
-
- Anderson, S. F., and J. E. Coleman. 1992. Conformational changes of HIV reverse transcriptase subunits on formation of the heterodimer: correlation with kcat and Km. Biochemistry 31:8221-8228. - PubMed
-
- Bardy, M., B. Gay, S. Pebernard, N. Chazal, M. Courcoul, R. Vigne, E. Decroly, and P. Boulanger. 2001. Interaction of human immunodeficiency virus type 1 Vif with Gag and Gag-Pol precursors: co-encapsidation and interference with viral protease-mediated Gag processing. J. Gen. Virol. 82:2719-2733. - PubMed
-
- Bathurst, I. C., L. K. Moen, M. A. Lujan, H. L. Gibson, P. H. Feucht, S. Pichuantes, C. S. Craik, D. V. Santi, and P. J. Barr. 1990. Characterization of the human immunodeficiency virus type-1 reverse transcriptase enzyme produced in yeast. Biochem. Biophys. Res. Commun. 171:589-595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
